Altingia excelsa
Altingia excelsa ist eine immergrüne Laubbaumart aus der kleinen Familie der Altingiaceae innerhalb der Ordnung der Steinbrechartigen (Saxifragales). Sie kommt in der östlichen Himalayaregion und in Südostasien vor. Unter dem Handelsnamen „Rasamala“ liefert sie ein wertvolles Nutzholz.
| Altingia excelsa | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
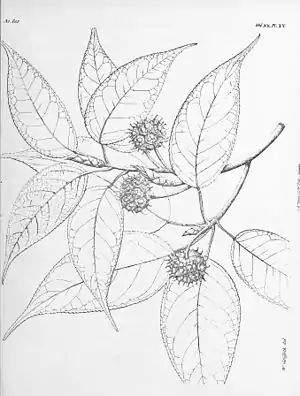
Altingia excelsa | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Altingia excelsa | ||||||||||||
| Noronha |
Beschreibung
Vegetative Merkmale
Der immergrüne Baum erreicht eine Höhe von 20–40(–50) m,[1] im Süden des Verbreitungsgebiets sogar 40–50(–60) m.[2] Dort kann der astfreie Schaft eine Länge von 20–35 m erreichen und der Stammdurchmesser 80–110(–200) cm betragen. Bäume, die freistehend kultiviert werden, haben dagegen einen niedrigen, rundlichen Habitus und haben nur einen schwach entwickelten Stamm.[2] Die Borke ist ziemlich glatt, hellgrau bis gelblich oder bräunlich-grau mit schmalen Längsrissen. Sie blättert leicht in unregelmäßigen, langen, dünnen Schuppen ab.
Die jungen Zweige sind kahl oder spärlich gelblich-braun flaumhaarig, ältere besitzen Lentizellen. Die mit Schuppen bedeckten Knospen sind schmal eiförmig.
Die schraubig angeordneten Laubblätter besitzen einen schlanken, (1,3–)2–3,5(–4) cm langen, kahlen bis spärlich gelblich-braun flaumhaarigen Stiel. Der Blattstiel weist für gewöhnlich an seinem oberen Ende einige sitzende oder kurz gestielte drüsenartige Fortsätze auf. Die einfache und ungeteilte, fiedernervige Blattspreite ist von elliptischer, länglicher oder eiförmiger bis eiförmig-lanzettlicher Form und hat eine Länge von 6–14(–17) cm und eine Breite von (2,5–)4–5,5(–7) cm. Sie besitzt einen breit keiligen oder abgerundeten bis fast herzförmigen Grund und ist vorne spitz bis zugespitzt, geschwänzt. Der Spreitenrand ist drüsig gekerbt-gesägt. Die dünn-ledrige, unterseits hellere Spreite ist oberseits kahl, unterseits minimal gelblich-braun flaumhaarig oder sehr selten samtig behaart. Auch bei sonst kahlen Spreitenunterseiten sind in den Nervenwinkeln Haarbüschel vorhanden. Die Spreite weist 5–8(–10) Paare von Seitennerven auf. Die hinfälligen, pfriemlichen Nebenblätter sind 1–6 mm lang und 0,25–0,8 mm breit.
Generative Merkmale
Die Geschlechtsverteilung der Blüten ist einhäusig gemischtgeschlechtig (monözisch). Die eingeschlechtlichen Blüten besitzen keine Blütenhülle. Sie sind jeweils getrennt in achsel- und/oder endständigen Köpfchen angeordnet.
Die männlichen Blütenstände sind vielblütige, ellipsoidale bis kugelige, 6–10 mm lange und 3–6 mm breite Köpfchen. Sie sind zu 6–14 zu ungefähr 2–3,5 cm langen, gelben „Trauben“ angeordnet. Die männlichen Blüten bestehen nur aus zahlreichen nicht miteinander verwachsenen Staubblättern mit basifixen, also an ihrem Grund dem dicken, 0,8–1 mm langen, kahlen Staubfaden angehefteten Staubbeuteln. Diese sind verkehrt-eiförmig und 1–1,4 mm lang. Die beiden oben gestutzten Theken bestehen aus jeweils zwei Pollensäcken und öffnen sich mit einem Schlitz der Länge nach. Zwischen den Staubblättern befinden sich einige linealische, 0,8–1 mm lange, mit winzigen feinen Flaumhaaren besetzte Blattorgane, die als Deckblätter gedeutet werden.
Die ähnlichen weiblichen Blütenstände erscheinen einzeln oder in traubigen Gruppen sowie unter den männlichen Gruppen. Es sind blass rosa gefärbte Köpfchen mit einem 2–4 cm langen, flaumhaarigen Stiel und sind am Grund von vier Hochblättern (Deckblätter) umgeben. Die Köpfchen haben einen Durchmesser von 5–9 mm und bestehen aus (4–)10–22 miteinander verwachsenen Blüten. Diese enthalten nur den halbunterständigen, zweikammerigen Stempel, der aus zwei miteinander verwachsenen, nur an Spitze freien Fruchtblättern besteht und einen ihn umgebenden, 0,3–0,5 mm hohen, gelappten Diskus sowie manchmal rudimentäre Staminodien. Die beiden zungenförmigen, 3–4 mm langen, auseinanderlaufenden und oft eingerollten, flaumhaarigen Griffel sind an der Innenseite gefurcht und besitzen eine herablaufende, papillöse Narbe. Jedes der beiden Fruchtknotenfächer enthält an der zentralwinkelständigen Plazenta zahlreiche Samenanlagen.
Die 20–38,5 mm lang gestielten,[2][3] holzigen Fruchtstände sind kugelig bis ellipsoid mit gestutzter Basis. Sie sind 14–20 mm lang, 15–25 mm breit und beinahe kahl. Die Fruchtstände enthalten (4–)9–25 Einzelfrüchte.[2][3] Dies sind holzige, zweifächerige Kapselfrüchte, die sich fachspaltig mit zwei zweiteiligen Klappen öffnen. Sie sind 8–10 mm lang, an der Spitze fein hellbraun flaumhaarig und von einem Ring von vergrößerten, verhärteten Diskuslappen umgeben. Der obere Teil der Griffel und die Staminodien sind im Fruchtzustand nicht mehr vorhanden. Die Samen sind in jedem Fruchtfach zahlreich vorhanden, aber die 15–25 oberen sind klein, unregelmäßig prismatisch geformt, ungeflügelt und steril. Immer nur höchstens einer der unteren Samen ist fruchtbar. Diese sind braun, verkehrt-eiförmig, abgeflacht und am Rand schmal geflügelt. Sie sind 4,4–6,1(–6,5) mm lang und 2,5–3,5 mm breit.[2][3] Der Nabel befindet sich zentral am Rücken des Samens. Die Samenschale ist dick und hart. Es ist nur wenig Endosperm vorhanden.
Altingia excelsa blüht hauptsächlich in den Monaten März bis Mai und kann das ganze Jahr über fruchten, hauptsächlich aber von Juni bis August. In Malesien gibt es auch noch eine zweite Blütezeit im Oktober und November.[2] Die Blüten erscheinen hier offenbar zusammen mit dem neuen Austrieb im Zuge des Laubwechsels beim Übergang von der Regenzeit zur Trockenzeit und umgekehrt. Die Art fruchtet hier dann schwerpunktmäßig in beiden Jahreszeiten jeweils in deren Mitte.
Chromosomen
Altingia excelsa hat einen diploiden Chromosomensatz mit 2n = 32.[4]
Verbreitung
Altingia excelsa ist im tropischen und subtropischen Asien in mehreren Teilarealen weit verbreitet. Die Art kommt in den Bergwäldern von Sumatra und West-Java sowie auf der Malaiischen Halbinsel in Malaysia vor. Ein weiteres Teilareal reicht von der östlichen Himalayaregion (Bhutan, Assam, Südost-Tibet) und Südost-Yunnan nach Myanmar, wo es im Süden bis nach Mergui reicht. Darüber hinaus wird Altingia excelsa im Zuge von Wiederaufforstungen angepflanzt.
Lebensraum und Ökologie
.jpg.webp)
Die Baumart wächst durchwegs in feuchten Bergwäldern in 550–1830 m Seehöhe.[1][2] Altingia excelsa tritt als strukturell dominierende Baumart auf, die das geschlossene Kronendach überragt. In Urwäldern können etwa 10–35 ausgewachsene Individuen pro Hektar vorhanden sein, deren Holzvolumen bis zu 150 m³ je Hektar ausmachen kann. Allerdings wurde die Baumart oft selektiv entnommen, sodass ungestörte Verhältnisse heute selten sind. In Malesien erreichen von den Begleitarten nur die Steineibengewächse Podocarpus neriifolius und Dacrycarpus imbricatus ähnliche Dimensionen. Weitere Begleitarten gehören hier zu den Gattungen Quercus, Schima, Castanopsis, Syzygium, Sloanea, Dysoxylum, Engelhardia, Magnolia und Elaeocarpus.[2]
Altingia excelsa wird seit 1851 auch wegen ihres wertvollen Holzes aufgeforstet, unter anderem auch in Zentral-Java, wo das Klima trockener ist als in ihrem natürlichen Verbreitungsgebiet. Damit das Wachstum zufriedenstellend ist, sollte der trockenste Monat des Jahres wenigstens 100 mm Niederschlag aufweisen.[5] Aufforstungen erfolgten zu Beginn mit Sämlingen aus Naturverjüngungen, später wurden dann auch Saatbeete angelegt. Die Samen sind nur für kurze Zeit keimfähig. Nach der Aussaat keimen sie schnell nach etwa einer Woche, aber die Sämlinge zeigen während der ersten paar Jahre nur geringes Wachstum. Erst danach folgt eine Periode mit raschem Zuwachs. Kultivierte Bäume zeigen das schnellste Wachstum in einer Höhenlage von 600–700 m.[5]
Gelegentlich wird Altingia excelsa völlig von Raupen entlaubt. Die Bäume treiben nach solchen Attacken rasch wieder aus. Zweierlei Gallen sind beobachtet worden, die eine verursacht von einer Gallmücke, die andere von einer Gallmilbe. Gallähnliche Zweigverdickungen können auch von der „Mistel“-Art Korthalsella japonica (= K. opuntia, Sandelholzgewächse) verursacht werden. Die Mistel Viscum stenocarpum (= V. liquidambaricolum auct.) kommt auf der Insel Java ausschließlich auf Altingia vor.[5]
Blüten- und Fruchtbiologie
Der Bau der Blüten ohne Schauapparat, mit großen Narben und staubförmigem Pollen macht es sehr wahrscheinlich, dass die Arten der Altingiaceae hauptsächlich durch den Wind bestäubt werden (Anemophilie).[6]
Die Samen sind ölhaltig. Sie werden deshalb von Affen und Vögeln gern gefressen. Insbesondere auch Ameisen ernten die Samen mit Vorliebe. Obwohl die Samen kein Elaiosom aufweisen, können sie daher trotzdem als myrmekochor angesehen werden. Die Ameisen bereiten in Baumschulen Probleme, wo sie Saatbeete mit Altingia excelsa in kurzer Zeit leerräumen können.[5]
Taxonomie
Die Art wurde 1790 vom spanischen Botaniker Francisco Noroña auf Basis eigener Aufsammlungen von der Insel Java beschrieben.[7] Es handelt sich dabei um eine posthume Veröffentlichung seiner Manuskripte. Altingia excelsa ist die Typusart der Gattung Altingia.
Sedgwickia cerasifolia Griff. und Liquidambar altingiana Blume sind Synonyme. Von manchen Autoren, beispielsweise in der „Flora of Thailand“,[8] wird auch Altingia siamensis Craib als Synonym zu Altingia excelsa gestellt. Nach R. Govaerts wird die Art am besten als Liquidambar excelsa (Noronha) Oken zu Liquidambar gestellt.[9]
Etymologie
Das Artepitheton excelsa (lat. hoch) leitet sich von lat. excellere (herausragen) her. Es bezieht sich auf die Wuchshöhe dieser Baumart.[10] Die Gattung Altingia ist zu Ehren von Willem Arnold Alting (1724–1800) benannt, dem Generalgouverneur von Niederländisch-Indien zur Zeit, als der Erstbeschreiber Francisco Noroña Java besuchte.[11]

Nutzung
Altingia excelsa ist unter dem Namen „Rasamala“[12] wegen seiner Tragkraft und Haltbarkeit eine der wertvollsten Nutzholzarten von West-Java.[13] Unter demselben Handelsnamen wird auch Holz aus Thailand, Laos, Kambodscha und Vietnam vermarktet,[12] das entsprechend seiner Herkunft eigentlich Altingia siamensis zuzuordnen ist. Das Holz wird in starken Stammdimensionen als Bauholz für Brückenkonstruktionen und Gebäude verwendet. Altes Kernholz wird nur langsam von Termiten befallen. Nachteile dieser Holzart sind das langsame Trocknen und eine Tendenz, sich zu werfen und zu reißen.[13]
Das Holz hat keine erkennbaren oder nur undeutliche Zuwachszonengrenzen. Das Kernholz ist braun bis rot gefärbt und ohne Farbstreifen, das Splintholz farblich davon deutlich abgesetzt. Die Rohdichte beträgt 0,6–0,85 kg/dm3.[12]
Die Borke enthält ein aromatisches Harz („Burmesisches Storax“[1]), das nach Verletzungen gebildet wird.[13] Es wird als Parfüm und als Räucherwerk verwandt.[1] Dies jedoch nur lokal und nicht in großem Stil.[13]
In West-Java werden junge Austriebe als Gemüse gegessen.[13]
Quellen
- D. G. Long: Family 65. Hamamelidaceae. In: A. J. C. Grierson, D. G. Long: Flora of Bhutan. Vol. 1, Nr. 3. Royal Botanic Garden, Edinburgh 1987, ISBN 0-9504270-6-3, S. 468–471.
- W. Vink: Hamamelidaceae. In: Flora Malesiana. Ser. I, Vol. 5, Nr. 3. Botanic Gardens of Indonesia, Bogor, Rijksherbarium, Leyden 1957, S. 363–379. (biodiversitylibrary.org, S. 376)
- Zhang Zhiyun, Zhang Hongda, P. K. Endress: Hamamelidaceae. In: Flora of China. Vol. 9: Pittosporaceae through Connaraceae. Science Press, Beijing, Missouri Botanical Garden Press, St. Louis 2003, ISBN 1-930723-14-8, S. 18–42. (efloras.org, Altingia excelsa)
Weblinks
Einzelnachweise
- D. G. Long 1987, S. 471.
- W. Vink 1957, S. 377. (biodiversitylibrary.org)
- S. M. Ickert-Bond, K. B. Pigg, J. Wen: Comparative infructescence morphology in Altingia (Altingiaceae) and discordance between morphological and molecular phylogenies. In: American Journal of Botany. Band 94, 2007, S. 1094–1115. doi:10.3732/ajb.94.7.1094
- Altingia excelsa bei Tropicos.org. In: IPCN Chromosome Reports. Missouri Botanical Garden, St. Louis, abgerufen am 2. Juni 2014.
- W. Vink 1957, S. 378. (biodiversitylibrary.org)
- P. K. Endress: Hamamelidaceae. In: K. Kubitzki, J. G. Rohwer, V. Bittrich (Hrsg.): The families and genera of Vascular Plants. Vol. II: Flowering Plants: Dicotyledons: Magnoliid, Hamamelid and Caryophyllid families. Springer, Berlin / Heidelberg / New York 1993, ISBN 3-540-55509-9, S. 322–331. (books.google.at), S. 324
- F. Noroña 1790: Altingia excelsa, malaice et javanice Rasamala, Lignum papuanum Rumphii herbar. Amboin., Vol. 2. Pag. 57. In: Verhandelingen van het Bataviaasch Genootschap der Kunsten en Weetenschappen. Band 5, Nr. 2, S. 1–9. (biodiversitylibrary.org)
- C. Phengklai: Hamamelidaceae. In: Flora of Thailand. Vol. 7, Nr. 3, 2001, The Forest Herbarium, Bangkok, S. 400–411.
- Rafaël Govaerts (Hrsg.): Liquidambar - World Checklist of Selected Plant Families des Royal Botanic Gardens, Kew. Zuletzt eingesehen am 14. September 2018.
- Helmut Genaust: Etymologisches Wörterbuch der botanischen Pflanzennamen. 3., vollständig überarbeitete und erweiterte Auflage. Birkhäuser, Basel / Boston / Berlin 1996, ISBN 3-7643-2390-6, S. 242.
- F. Noroña 1790, S. 6. (biodiversitylibrary.org)
- H. G. Richter, M. J. Dallwitz: Altingia excelsa. In: Handelshölzer. Abgerufen am 7. Juli 2014.
- W. Vink 1957, S. 379. (biodiversitylibrary.org)