Ökosystem See
Das Ökosystem See gehört zu den limnischen Ökosystemen, die von stehenden Gewässern gebildet werden.
Die folgenden Betrachtungen beziehen sich auf einen Süßwassersee in der gemäßigten Klimazone, ein stehendes Gewässer ohne direkten Abfluss zum Meer, das mehr als 10 Meter tief ist und dessen Verdunstungsrate sowie Zu- und Abflussmenge gegenüber der gesamten Wassermenge gering ist (vgl. Gewässer#Gewässertypen).
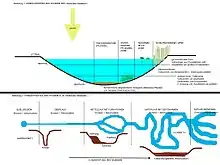
Vertikale Gliederung

Die Freiwasserzone (Pelagial) wird untergliedert in:
- Trophogene Zone (Nährschicht): In den oberflächennahen Wasserschichten ist genügend Licht für die Photosynthese der Primärproduzenten (Wasserpflanzen, Algen und vor allem Phytoplankton) vorhanden. Diese produzieren durch die Photosynthese mehr Sauerstoff und Biomasse, als durch die eigene Zellatmung und die Atmung der heterotrophen, aeroben Organismen (Zooplankton, Destruenten und Nekton) verbraucht werden. Die Lichtintensität nimmt aber mit der Tiefe annähernd exponentiell ab, abhängig von der Dichte der "Trübstoffe", insbesondere der Algen und Zooplankter.
- Kompensationsebene: Die Kompensationsebene bildet die Grenze zwischen der trophogenen und der tropholytischen Zone. Sie liegt ungefähr in der Tiefe, in der die Photosynthetisch aktive Strahlung noch eine Restintensität von 1 % des Wertes dessen direkt unter der Oberfläche hat. Nur oberhalb dieser Grenze ist eine positive Energiebilanz zwischen Photosynthese und Zellatmung (positive Nettoproduktion) möglich. Die Tiefe der Ebene ist dynamisch: Photoautotrophe Organismen haben artspezifische Kompensationstiefen und die Tiefe schwankt mit den optischen Eigenschaften des Wassers und den aktuellen Lichtverhältnissen.[1]
- Tropholytische Zone (Zehrschicht): Unterhalb der Kompensationstiefe ist fast keine Photosynthese mehr möglich. Die heterotrophen Organismen verbrauchen Sauerstoff und Biomasse, die aus der trophogenen Zone nach unten absinkt. Absinkende Algen veratmen ihre Vorräte und sterben ab.

Das Pelagial kann auch mittels thermischer Analyse zoniert werden. Folgende Gliederung des Sees in verschiedene Tiefenzonen bildet sich während der sogenannten Sommerstagnation durch die Temperaturschichtung aus. Während der Zirkulationsphasen im Frühjahr und Herbst wird diese Schichtung wieder beseitigt.
- Epilimnion (Oberflächenwasser): Die Temperatur unterliegt relativ großen tages- und jahreszeitlichen Schwankungen zwischen 4 °C und über 30 °C, abhängig von der Solarstrahlung, der Lufttemperatur und den Windverhältnissen. Der Sauerstoffgehalt ist hoch, da ständig Sauerstoff ins Wasser diffundiert. Der Sauerstoffgehalt ist weiterhin abhängig von der Wassertemperatur, der Durchmischung mit der Luft durch Wind und Wellen und von den vorherrschenden Organismen. Während der Stagnation wird nur das Epilimnion vom Wind durchmischt und das Metalimnion bleibt nährstoffarm, was die Primärproduktion entscheidend begrenzt.[2][3]
- Metalimnion (Sprungschicht): Das Metalimnion ist gekennzeichnet durch eine rasche Temperaturabnahme (steiler Temperaturgradient) mit der Tiefe auf 4 °C (Thermokline, Temperatursprungschicht). Der Sauerstoffgehalt nimmt rasch ab, abhängig von der Zahl der aeroben Lebewesen.
- Hypolimnion (Tiefenschicht): In dieser Schicht herrscht eine tages- und jahreszeitlich unabhängige, mehrheitlich konstante Temperatur von 4 °C und eine relativ niedrige Sauerstoffkonzentration. Die Schicht ist bedingt durch die Dichteanomalie des Wassers, aufgrund der die Dichte von Wasser bei 3,98 °C am höchsten ist. Ferner wird die Dichte noch vom Druck und der Menge an gelösten Stoffen beeinflusst.[4]

Die Bodenzone (Benthal – gr. benthos, tief) wird ebenfalls durch die Kompensationsebene unterteilt:
- Litoral – Uferzone (lat. litus, litoris, Ufer): Das Litoral umfasst den Uferbereich bis zur Kompensationstiefe. Es wird überwiegend von höheren Pflanzen bewachsen.
- Profundal – Tiefenzone (lat. profundus, tiefgründig): Im Profundal leben keine photoautotrophen Produzenten. Konsumenten sind demnach von der Biomasse abhängig, die das Litoral und die trophogene Zone bildet.
Litoral
Das Litoral wird in einzelne Zonen oder Gürtel aufgrund der Lebensgemeinschaften unterteilt:
- Epilitoral, Erlengürtel, Weidengebüschzone
- Das Epilitoral ist der oberste Rand des Ufers, der nicht mehr vom Wellenschlag betroffen ist. Aufgrund des hoch anstehenden Grundwasserspiegels stellt er einen Lebensraum für Pflanzen dar, die mit einem feuchten Boden zurechtkommen (Hygrophyten). Die lichten Sumpf- und Bruchwälder mit Schwarzerle (Alnus glutinosa), Moorbirke (Betula pubescens) und Weidenarten (Salix spp.) haben einen dichten Unterwuchs, der unter anderem aus Moosen, Farnen, Sauer- oder Riedgräsern und Binsen besteht. Daneben findet man auch die Sumpf-Schwertlilie (Iris pseudacorus) und die Sumpfdotterblume (Caltha palustris).
- Die Tierwelt setzt sich zunächst aus Detritusfressern (Regenwürmer, z. B. Lumbricus terrestris, Asseln, Landschnecken), Spinnen und Insekten zusammen, die wiederum die Nahrungsgrundlage für Vögel darstellen, die Bäume, Büsche und den Boden als geschütztes Brutrevier nutzen.
- Supralitoral, Spritzwasserzone
- Dieser Uferstreifen wird zwar von den Wellen selbst nicht erreicht, aber vom Spritzwasser der sich am Ufer brechenden Wellen durchnässt.
- Eulitoral, Brandungszone
- In der Brandungszone herrschen starke mechanische Kräfte, die das Aufwachsen größerer Pflanzen nicht zulassen. Jedoch siedeln sich fest haftende, sauerstoffliebende Organismen wie Strudelwürmer (Turbellaria spec.) sowie krustenbildende Cyanobakterien hier an.
- Infralitoral oder Sublitoral
- In dieser Zone siedeln größere Pflanzen, die an einen ständig überfluteten Boden angepasst sind. Sie besitzen ein Aerenchym, ein zusammenhängendes System von luftgefüllten großen Zellzwischenräumen (Interzellulare), so dass auch die Wurzeln mit Sauerstoff versorgt werden können. Dieser Uferbereich dient als Laich- und Brutgebiet für viele Fische, Vögel und Insekten. Das Infralitoral wird in mehrere Abschnitte unterteilt:
- Großseggenzone
- Sie liegt im Bereich zwischen Hoch- und Niedrigwassermarke. Charakteristische Pflanzen sind die Seggen (Carex spp.). Daneben kommen auch Wasserminze (Mentha aquatica), Blutweiderich (Lythrum salicaria) und Sumpf-Schachtelhalm (Equisetum palustre) vor.
- Röhrichtzone
- Auch hier sind noch emerse Pflanzen, deren Stängel und Blätter größtenteils über den Wasserspiegel hinausragen, zu finden. Dies sind vor allem Schilfrohr (Phragmites australis), Rohrkolben (Typha sp.) und Froschlöffel (Alisma sp.). Blässhuhn (Fulica atra) und Teichralle (Gallinula chloropus) nisten hier.
- Schwimmblattzone
- In windgeschützten Teilen des Sees können sich Schwimmblattpflanzen ansiedeln, die bis auf die Blätter vollständig untergetaucht sind. Diese Blätter schwimmen auf der Wasseroberfläche und haben die Spaltöffnungen für den Gasaustausch auf der Oberseite der Blätter. (Bei Landpflanzen befinden sich die Stomata in der Regel auf der Blattunterseite). Am auffälligsten sind neben dem Wasser-Knöterich (Persicaria amphibia) die Vertreter der Seerosengewächse, Seerose (Nymphaea alba) und Teichrose (Nuphar lutea).
- Laichkrautzone
- Hier leben nahezu vollständig untergetauchte Wasserpflanzen, die auch unterhalb der Wasserlinie Blätter aufweisen, die dann oft stark zergliedert sind, um die Oberfläche für den Stoffaustausch zu vergrößern. Namengebend sind die Laichkräuter (Potamogeton spp.).
- Characeenzone
- Die Pflanzen dieser Zone sind vollständig untergetaucht. Blütenpflanzen wie das wurzellose Hornblatt (Ceratophyllum sp.), Tausendblatt (Myriophyllum sp.) oder Wasserpest (Elodea canadensis) können aufgrund zu geringer Lichtstärke nicht in eine Tiefe über 10 m vordringen. Wenn das Licht ausreicht, können aber die genügsameren Moose, zum Beispiel das Quellmoos (Fontinalis antipyretica) und bodenlebenden (benthischen) Algen bis zu 30 Meter Tiefe vorkommen. So bilden die Armleuchteralgen (Characeae) die unterste Zone der unterseeischen Wiesen.
Die einzelnen Zonen des Litorals bilden für Tiere unterschiedliche ökologische Nischen, die es ihnen ermöglichen, trotz ähnlicher Nahrungsansprüche Konkurrenz zu vermeiden.
Beispiele:
- Die Stockente (Anas platyrhynchos) ist bei der Nahrungssuche nur in seichteren Uferzonen zu finden, da sie beim Gründeln nicht abtaucht. Der Höckerschwan (Cygnus olor) kann mit seinem längeren Hals den Boden in tieferem Wasser nach Nahrung durchsuchen, während der Haubentaucher (Podiceps cristatus) in größeren Tiefen Jagd nach Fischen macht.
- Der Nachtreiher (Nycticorax nycticorax) findet seine Nahrung (Kleinsäuger, Amphibien, Insekten, Würmer) im Epilitoral, der Purpurreiher (Ardea purpurea) mit einem ähnlichen Nahrungsspektrum geht in der Röhrichtzone auf Jagd.
Freiwasserzone (Pelagial)
In der lichtdurchflutenden Freiwasserzone findet man vor allem Phytoplankton, auf der Oberfläche zuweilen auch frei schwimmende Pflanzen (Neuston und Pleuston) wie Wasserlinsen (Lemna spec.) oder den Schwimmfarn (Salvinia natans).
Die gesamte Freiwasserzone ist Lebensraum für Zooplankton, Nekton und Destruenten.
Die wichtigsten Plankter eines europäischen Süßwasser-Sees (Beispiel: Bodensee).
- Phytoplankton: Goldalgen (Dinobryon), Rhodomonas, Diatomeen (Tabellaria, Diatoma, Fragilaria, Asterionella), Grünalgen (Chlorella, Scenedesmus)
- Zooplankton: Ruderfußkrebse (Eudiaptomus, Cyclops, Mesocyclops), Blattfußkrebse (Daphnia), Rädertierchen
Zirkulation
In einem See kommt es regelmäßig zu einer teilweisen oder vollständigen Durchmischung (Zirkulation) des Wasserkörpers, wobei Sauerstoff und Nährstoffe über den durchmischten Bereich gleichmäßig verteilt werden. Die Antriebskräfte für die Zirkulationen sind Wind und Dichteunterschiede (kaltes, also dichteres Wasser sinkt nach unten, warmes, also weniger dichtes, steigt auf). Die Zirkulation kann weiterhin durch Dichteunterschiede des Wassers (auf Grund von Temperatur- oder Salzgehaltsunterschieden) begünstigt oder behindert werden. Hat sich eine Thermokline (ein Temperatursprung) ausgebildet, wirkt das Metalimnion wie eine Sperrschicht, das kalte, dichte Tiefenwasser kann nicht mehr an der Zirkulation teilnehmen, es findet nur eine Teilzirkulation und damit nur eine Durchmischung des Epilimnions statt. Dieser Zustand wird als Sommerstagnation bezeichnet.
Dies hat Folgen für die Sauerstoffversorgung der aeroben Organismen des Hypolimnions und für die Nährsalz- und Kohlenstoffdioxidversorgung der Primärproduzenten des Epilimnions:
Zwar erhalten die aeroben Organismen des Hypolimnions noch weiterhin genügend Nährstoffe, wenn abgestorbene Körper der Tiere und Pflanzen und anderes organisches Material (Detritus) nach unten sinken. Die Versorgung mit Sauerstoff aus dem Epilimnion ist aber unterbrochen. Das Hypolimnion verarmt verstärkt an Sauerstoff.
Die aeroben Destruenten des Hypolimnions, vor allem in der Bodenzone, remineralisieren das organische Material, es entstehen vor allem in Wasser lösliche Nitrate und Phosphate, die als Nährsalze für die Produzenten notwendig wären, um Photosynthese zu betreiben. Aufgrund des fehlenden Austauschs mit dem Epilimnion können diese Nährsalze und das bei der Dissimilation entstehende Kohlenstoffdioxid aber nicht ins Epilimnion gelangen.
Deshalb beobachtet man nach der Ausbildung einer Sprungschicht einen Rückgang der Phytoplanktonmasse im Epilimnion.
Bricht aufgrund der Abkühlung des Oberflächenwassers die Thermokline zusammen, ist wieder Vollzirkulation möglich. Nährsalze und Kohlenstoffdioxid gelangen ins Epilimnion, was zu einer Vermehrung der Produzenten führt. Bei sehr starker Vermehrung des Phytoplanktons spricht man dann von Algenblüte.
Klassifikation von Seen nach der Anzahl der Vollzirkulationen im Jahr
- Amiktisch: Arktische und antarktische Gewässer sind von Dauereis bedeckt, das eine Zirkulation vollständig unterbindet.
- Monomiktisch: Eine Vollzirkulation findet nur in einer Jahreszeit statt:
- Polare und subpolare Seen: Vollzirkulation während der Sommermonate; Winterstagnation bei Eisbedeckung.
- Subtropische Seen: Vollzirkulation im Winter bei ausreichender Abkühlung des Oberflächenwassers, Stagnation im Sommer. Beispiele: Lago Maggiore, Gardasee, Bodensee als nördlichster dieses Typs.
- Dimiktisch: Vollzirkulation im Frühjahr und im Herbst, in temperierten Seen der gemäßigten Breiten.
- Frühjahrszirkulation (Vollzirkulation): Im Frühjahr erwärmt sich das Oberflächenwasser. Frühjahrsstürme sorgen für eine vollständige Durchmischung des Sees.
- Sommerstagnation (Teilzirkulation): Im Sommer erwärmt sich das Oberflächenwasser deutlich stärker als das Tiefenwasser. Es bildet sich ein deutlicher Temperaturgradient aus, der den Bereich des Metalimnions kennzeichnet. Darunter liegt bei ausreichender Seetiefe ein bei 4 °C homogener Bereich, das Hypolimnion. Die ständigen Zirkulationen durch Wind und nächtliche Konvektion beschränken sich auf das sich dadurch ausbildende Epilimnion, dessen Tiefe je nach Wetterlage schwankt.
- Herbstzirkulation (Vollzirkulation): Im Herbst kühlt das Oberflächenwasser ab, verdichtet sich und sinkt ab. Mit ihm senkt sich auch die zunehmend engräumigere Temperatursprungschicht. Unterstützt durch die Herbststürme kommt es zur Vollzirkulation.
- Winterstagnation (keine Zirkulation): Im Winter sinkt die Temperatur des Oberflächenwassers unter 4 °C und verliert damit an Dichte. Es entwickelt sich eine instabile inverse Temperaturverteilung (unter 4 °C kaltes Oberflächenwasser über 4 °C kaltem Tiefenwasser). Wenn Eis die Seeoberfläche bedeckt, wird die Temperaturschichtung stabilisiert.
- Oligomiktisch: Bei tropischen Tieflandseen wird das Oberflächenwasser stark erwärmt, nur selten durchbrechen unregelmäßige Vollzirkulationen die Dauerstagnation.
- Polymiktisch
- tropische Hochgebirgsseen: Sie weisen das ganze Jahr hindurch anhaltende, durch den Wind und nächtliche Abkühlung verursachte Vollzirkulation auf. Beispiel: Titicacasee in 3.810 m Höhe
- Seen im Hügelland der Tropen: Die starke Erwärmung am Tag führt zu einer Stagnation, die starke nächtliche Abkühlung zur Vollzirkulation.
- Flachseen der gemäßigten Breiten, wo es aufgrund der geringen Wassertiefe zu keiner Schichtung kommen kann. Das Wasser zirkuliert, durch den Wind angetrieben, ganzjährig. Wegen der Dichteanomalie des Wassers treten bei geschlossener Eisdecke jedoch Stagnationsphasen auf.
Klassifizierung nach der Reichweite der Vollzirkulation
- Holomiktisch (gr. holos = ganz) Die Vollzirkulation erfasst alle Wassermassen des Sees. Bei Flachgewässern der mittleren Breiten erfolgt die Wasserdurchmischung durch Konvektion und Wind.
- Meromiktisch (gr. meros = Teil): Die Vollzirkulation ist nicht bis zum Seegrund möglich. Die bei der Vollzirkulation nicht erfassten Wassermassen heißen Monimolimnion. Gründe können hierfür sein:
- eine windgeschützte Lage des Sees: etliche Kärntner Seen.
- Die Wasseroberfläche bietet im Verhältnis zur Tiefe eine zu kleine Angriffsfläche für den Wind. Beispiel: Königssee
- Es kommt zur Ausbildung einer Salzsprungschicht (Halokline), da das salzreichere und damit dichtere Wasser in die Tiefe sinkt. Dies geschieht, wenn ein See eine relativ hohe Salzfracht aufgrund seiner Zuflüsse und einer hohen Verdunstungsrate hat.
Nährstoffe
Der Gehalt an Nährsalzen, vor allem Phosphaten und Nitraten sowie an organischen Nährstoffen bestimmt, welche Arten, wie viel verschiedene Arten und wie viele Individuen in einem See leben können.
Der Nährstoffgehalt eines Sees wirkt sich vor allem während der Sommerstagnation aus:
Auswirkungen des Nährstoffgehaltes
Je mehr Nährsalze im Frühjahr durch die Vollzirkulation ins Epilimnion gelangen, umso mehr Phytoplankton (Schwebalgen) kann wachsen. Davon abhängig vermehren sich auch die Konsumenten. Bei eutrophen Seen wird durch das hohe Angebot an Nährstoffen, insbesondere Phosphat, die Biomasse so groß, dass es zur sog. Algenblüte kommen kann.
Beispiel: Änderung abiotischer Faktoren im Jahresverlauf im Epilimnion eines dimiktischen eutrophen Sees in Europa
 |
 |
| Das Sauerstoffmaximum im Mai korrespondiert mit dem Absinken der übrigen Faktoren: Aufgrund der steigenden Temperaturen und der verbesserten Lichtverhältnisse sowie des hohen Nährsalzangebotes durch die Vollzirkulation im Frühjahr kommt es zu einer Massenvermehrung der Fotosynthese betreibenden Organismen. Da dies dazu führt, dass in den unteren Schichten des Epilimnions nicht mehr genügend Licht vorhanden ist, stirbt dort das Phytoplankton ab und sinkt zu Boden. | Der Anstieg von Kohlenstoffdioxid im März und November erfolgt aufgrund der Frühjahrs- und Herbst-Vollzirkulation. Die Bildung von Kohlenstoffdioxid durch die Atmung der Konsumenten wird während der Sommerstagnation von dem Verbrauch durch die Photosynthese überkompensiert. |
 |
 |
| Der Anstieg des Phosphatgehaltes erfolgt im März und November aufgrund der Frühjahrs- und Herbst-Vollzirkulation. Eine Massenvermehrung der Produzenten im April und Mai verbraucht nahezu das ganze Phosphat. | Der Anstieg des Nitratgehaltes im März und November erfolgt aufgrund der Frühjahrs- und Herbst-Vollzirkulation. Eine Massenvermehrung der Produzenten im April und Mai verbraucht sehr viel Nitrat, das aber durch die Fixierung von Luftstickstoff von Cyanobakterien teilweise ersetzt wird. |
Einteilung nach der Nährstoffmenge
 |
- Oligotrophe (gr. oligos = wenig; gr. trophē = Nahrung) Seen enthalten nur wenig Nährsalze; dadurch ist die Vermehrung des Phytoplanktons und der Konsumenten begrenzt. Da nur wenig totes, organisches Material anfällt, sind diese Seen das ganze Jahr hindurch in allen Wassertiefen gut mit Sauerstoff versorgt. Aufgrund der geringen Dichte des Phytoplanktons ist die Sichttiefe groß (bis zu 10 m), das Wasser erscheint klar, der See hat eine blaue oder grüne Farbe. Am Boden sammelt sich kein Faulschlamm, da für die aeroben Destruenten genügend Sauerstoff vorhanden ist, um das organische Material rasch abzubauen. Oligotrophe Seen zeichnen sich durch einen großen Artenreichtum aus. Es handelt sich hierbei zumeist um Stenoxybionten[5], die nur geringe Sauerstoffschwankungen tolerieren. Gelegentlicher Stickstoffmangel wird durch Stickstoff-fixierende, photoautotrophe Cyanobakterien ausgeglichen. Die geringe, meist organisch gebundene Phosphatmenge stammt vorwiegend aus Zuflüssen. Die in sauerstoffreichem Wasser vorhandenen freien Eisen(III)-Ionen (Fe3+) binden das Phosphat als schwerlösliches FePO4 und entziehen es dem Stoffkreislauf des Sees. Die Uferzone ist schmal und mit wenigen Makrophyten bewachsen. Ungestörte, subalpine Seen sind in der Regel oligotroph (Beispiel: Königssee).
- Mesotrophe (gr. mesos = der Mittlere) Seen enthalten mehr Nährsalze und damit eine größere Menge an Produzenten, Konsumenten und Destruenten. Der Sauerstoffgehalt im Hypolimnion während der Sommerstagnation ist zeitweilig gering.
- Eutrophe (gr. eu = gut, gediegen) Seen enthalten viele Nährsalze. Dadurch kommt es nach den Vollzirkulationen im Frühjahr und im Herbst zu einer starken Vermehrung von Phytoplankton und Konsumenten. Lichtmangel und Nährsalzmangel lassen die Algen aber bald wieder absterben, so dass die Biomasse im Epilimnion während der Sommerstagnation wieder geringer ist. Da nun sehr viel totes, organisches Material anfällt, kommt es zu einem Sauerstoffdefizit im Hypolimnion. Aufgrund der hohen Dichte des Phytoplanktons ist die Sichttiefe gering, das Wasser erscheint trübe, der See hat eine braungrüne Farbe. Am Boden sammelt sich eine dicke, aus organischem Plankton-Detritus bestehende, Halbfaulschlammschicht (Gyttja), wenn die aeroben Destruenten aufgrund des Sauerstoffmangels durch anaerobe ersetzt werden. Während der Vollzirkulationen kann diese Faulschlammschicht durch aerobe Destruenten wieder weitgehend abgebaut werden. Reicht der Sauerstoff hierfür nicht, kommt es zum Umkippen des Sees durch die Entstehung einer positiven Rückkopplung zwischen der Dauerlöslichkeit von Phosphat (mit Fe2+) und der erhöhten Primärproduktion. Dies ist dann der Übergang zum hypertrophen Zustand (siehe unten). Eutrophe Seen zeichnen sich durch einen geringeren Artenreichtum, aber durch eine hohe Individuendichte aus. Es handelt sich hierbei zumeist um Euroxybionten[6], welche die zeitlichen und räumlichen starken Sauerstoffschwankungen tolerieren. Die Uferzone ist breit und dicht bewachsen.
- Hypertrophe (gr. hypér = groß, übermäßig) Seen enthalten sehr viel totes organisches Material, das sich am Boden als zuweilen mehrere Meter mächtige Faulschlammschicht ablagert. Das Hypolimnion enthält das ganze Jahr über kaum Sauerstoff. Statt aerobe bauen anaerobe Destruenten das organische Material ab. Dieser Abbau erfolgt aber sehr viel langsamer und unvollständig. Außerdem entstehen statt Kohlenstoffdioxid und Mineralsalzen wie Nitrat und Sulfat giftige Stoffe wie Methan, Ammoniak und Schwefelwasserstoff. Diese Stoffe und der Sauerstoffmangel machen den See zu einem instabilen Gewässer, in dem nur wenige Arten, diese aber in z. T. gewaltigen Individuenzahlen, leben können. Diese Arten sind zwar an Sauerstoffmangelbedingungen mehr oder weniger angepasst, aber dennoch ständig von katastrophalen Zusammenbrüchen der Bestände bei ungünstigen Bedingungen bedroht. Hypertrophe Seen verlanden vom Ufer aus, da sich anfallender Detritus im Pflanzendickicht des Litorals schneller ansammelt.
Nahrungsnetz
Ein Nahrungsnetz stellt im Idealfall alle Nahrungsbeziehungen zwischen den Organismen eines Ökosystems dar. Um die Übersichtlichkeit zu wahren, ist im folgenden Beispiel nur ein Ausschnitt aus dem Nahrungsnetz eines europäischen Sees dargestellt. Auf Aasfresser und Destruenten wurde dabei verzichtet.
 |
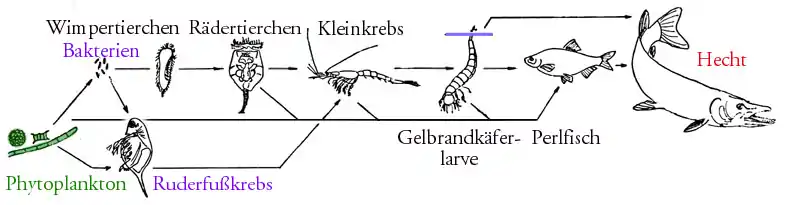 Nahrungsnetz-Ausschnitt (zu Wasserflöhe (Daphnia), Wimpertierchen (Paramecium), Rädertierchen (Brachionus), Kleinkrebs (Mysis), Gelbrandkäferlarve (Dytiscus), "Perlfisch" auf dem Bild ist eine Rotfeder (Scardinius) abgebildet und Hecht (Esox) siehe dort) |
Nahrungsketten im Pelagial (Freiwasserbereich in einem See):
| Trophieebene | |
|---|---|
| Phytoplankton | Produzent |
| herbivore Zooplankter | Primärkonsument |
| carnivore Zooplankter | Sekundärkonsument |
| pelagische Friedfische | Tertiärkonsument |
| pelagische Raubfische | Endkonsument |
| aerobe und anaerobe Bakterien | Destruenten |
Globale Bedeutung
Die Bruttoprimärproduktion (GPP, gross primary production) aller Seen weltweit ist auf 0,65 Petagramm Kohlenstoff pro Jahr abgeschätzt worden[7] (1 Petagramm = 1 Milliarde Tonnen). Das ist im Verhältnis zur gesamten Bruttoprimärproduktion von 100 bis 150 Pg C/Jahr nicht viel. Seen sind darüber hinaus offensichtlich Netto-Quellen, und nicht etwa Senken, von Kohlendioxid[8][9]. Gleichzeitig werden erhebliche Mengen Kohlenstoff in Seensedimenten festgelegt und so dem Kohlenstoffkreislauf entzogen[10]. Dieser scheinbare Widerspruch löst sich auf durch den erheblichen Zustrom von Biomasse aus terrestrischen Ökosystemen in Seen hinein. In Seen, die von Karbonatgesteinen umgeben sind, spielt auch der Einstrom in Form von gelöstem (Hydrogen-)Carbonat eine große Rolle, die diejenige der seeneigenen Atmung (Respiration) sogar übersteigen kann. Seen sind eben nur bei idealisierter Betrachtung isolierte Ökosysteme, in der Realität sind sie mit den terrestrischen Systemen eng gekoppelt und verwoben. Auch der organische Kohlenstoffzustrom in Seen erfolgt weit überwiegend in gelöster Form, nur ganz untergeordnet in fester. Die globale Nettoemission von Seen wird in der Übersichtsarbeit von Tranvik et al. auf 0,53 Petagramm Kohlenstoff pro Jahr abgeschätzt. Das ist auch global betrachtet ein signifikanter Wert, der erstaunt, wenn man bedenkt, dass weniger als 3 % der kontinentalen Fläche von Seen eingenommen wird; der Wert ist mehr als halb so hoch wie der Nettoexport von Kohlendioxid in die Ozeane. Seen, insbesondere eutrophe Flachwasserseen, tragen auch zur Methanproduktion bei, ihr Anteil wird aber auf lediglich auf 6 bis 16 % der natürlichen Gesamtemissionen (d. h. ohne menschliche Quellen) geschätzt. Auch die Kohlenstoffspeicherung in Seensedimenten ist beachtlich. Der globale Vorrat an organisch gebundenem Kohlenstoff wurde auf 820 Petagramm geschätzt, das wäre das dreifache des in den Ozeansedimenten gespeicherten. Man muss dabei aber beachten, dass die gesamte Tiefsee mit ihren Sedimenten zum Kohlenstoffspeicher überhaupt nichts beiträgt. Die jährliche Nettospeicherrate der Seensedimente könnte etwa 0,03 bis 0,07 Pg C betragen[11]. Je nach spezieller Ausgestaltung des Ökosystems können Seen also sowohl Quellen wie auch Senken für Kohlendioxid sein. Die Auswirkungen künftiger Veränderungen vorherzusagen, ist wegen der zahlreichen Prozesse mit zueinander gegenläufigen Ergebnissen sehr schwierig.
Siehe auch
Literatur
- Jürgen Schwoerbel et al.: Einführung in die Limnologie. 9. Auflage. Elsevier GmbH, München 2005, ISBN 3-8274-1498-9.
- Eberhard Schmidt: Ökosystem See. Der Uferbereich des Sees. 5. Auflage. Quelle & Meyer Verlag GmbH & Co., Wiesbaden 1995, ISBN 3-494-01152-4.
Einzelnachweise
- Jürgen Schwoerbel et al.: Einführung in die Limnologie. 9. Auflage. Elsevier GmbH, München 2005, ISBN 3-8274-1498-9, S. 54, 130, 131.
- Thomas M. Smith et al.: Ökologie. 6. Auflage. Pearson Education Deutschland GmbH, München 2009, ISBN 978-3-8273-7313-7, S. 616, 617.
- Jürgen Schwoerbel et al.: Einführung in die Limnologie. 9. Auflage. Elsevier GmbH, München 2005, ISBN 3-8274-1498-9, S. 37.
- Jürgen Schwoerbel et al.: Einführung in die Limnologie. 9. Auflage. Elsevier GmbH, München 2005, ISBN 3-8274-1498-9, S. 27.
- Arten, deren Stoffwechselrate etwa linear vom Sauerstoffgehalt im Umgebungsmedium abhängt; sie benötigen daher eine Umgebung mit hohem Sauerstoffgehalt. definiert in: Pierre Lasserre: Metabolic Activities of Benthic Microfauna and Meiofauna. In I. N. McCave (Hrsg.): The Benthic Boundary Layer. Springer Verlag, 1976, ISBN 978-1-4615-8749-1, Definition auf S. 109.
- Arten, die ihre Stoffwechselrate regulieren und an den Sauerstoffgehalt des Umgebungsmediums anpassen können; sie können daher eine Umgebung mit niedrigem Sauerstoffgehalt tolerieren. definiert in: Pierre Lasserre: Metabolic Activities of Benthic Microfauna and Meiofauna. In I. N. McCave (Hrsg.): The Benthic Boundary Layer. Springer Verlag, 1976, ISBN 978-1-4615-8749-1, Definition auf S. 109.
- M.L. Pace & Y.T. Prairie (2005): Respiration in lakes. In: P.A. del Giorgio & P.J.L.B. Williams (Hrsg.): Respiration in aquatic ecosystems. Oxford Univ. Press. pp. 103–122.
- Jonathan J. Cole, Nina F. Caraco, George W. Kling, Timothy K. Kratz (1994): Carbon Dioxide Supersaturation in the Surface Waters of Lakes. Science Vol.265 no.5178: 1568-1570 doi:10.1126/science.265.5178.1568
- Lars J. Tranvik, John A. Downing, James B. Cotner, Steven A. Loiselle, Robert G. Striegl, Thomas J. Ballatore, Peter Dillon, Kerri Finlay, Kenneth Fortino, Lesley B. Knoll, Pirkko L. Kortelainen, Tiit Kutser, Soren Larsen, Isabelle Laurion, Dina M. Leech, S. Leigh McCallister, Diane M. McKnight, John M. Melack, Erin Overholt, Jason A. Porter, Yves Prairie, William H. Renwick, Fabio Roland, Bradford S. Sherman, David W. Schindler, Sebastian Sobek, Alain Tremblay, Michael J. Vanni Antonie M. Verschoor, Eddie von Wachenfeldt, Gesa A. Weyhenmeyer (2009): Lakes and reservoirs as regulators of carbon cycling and climate. Limnology and Oceanography 54(6, part 2): 2298–2314.
- Walter E. Dean & Eville Gorham (1998): Magnitude and Significance of Carbon Burial in Lakes, Reservoirs, and Peatlands. Geology v. 26; no. 6: 535–538. download
- J.J. Cole, Y.T. Prairie, N.F. Caraco, W.H. McDowell, L.J. Tranvik, R.G. Striegl, C.M. Duarte, P. Kortelainen, J.A. Downing, J.J. Middelburg, J. Melack (2007): Plumbing the Global Carbon Cycle: Integrating Inland Waters into the Terrestrial Carbon Budget. Ecosystems 10: 171–184. doi:10.1007/s10021-006-9013-8