Qlikogen
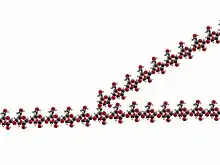
Qlikogen — Qlikogen, heyvanlarda, göbələklərdə və bakteriyalarda enerji saxlama forması kimi xidmət edən çox dallı bir qlükoza polisaxariddir. Polisaxaridin quruluşu bədəndə qlükoza saxlamağın əsas formasıdır[2].

.tif.jpg.webp)
Qlikogen iki enerji ehtiyatı formasından biri kimi fəaliyyət göstərir: qlikogen qısa müddətli istifadə üçün, digər forması isə uzun müddət saxlanması üçün yağ toxumasında olan triqliserid anbarlarıdır. İnsanlarda qlikogen əsasən qaraciyər və skelet əzələlərinin hüceyrələrində istehsal olunur və saxlanılır. Qaraciyərdə qlikogen orqanizmin yaş çəkisinin 5–6% -ni, 1,5 kq ağırlığında bir yetkinin qaraciyəri isə təxminən 100–120 qram glikogen saxlaya bilir. Skelet əzələlərində aşağı konsentrasiyalarda qlikogen (əzələ kütləsinin 1–2% -i), 70 kq ağırlığında bir yetkinin skelet əzələlərində təxminən 400 qram qlikogen saxlanılır[3].
Təxminən 4 qram qlükoza qanda daimi olaraq mövcuddur. Ac insanlarda qaraciyər və skelet əzələlərindəki qlikogen depoları tərəfindən qan qlükoza səviyyələri bu səviyyədə sabit saxlanılır. Skelet əzələlərinin qlikogen anbarları əzələlərin özləri üçün enerji saxlama formasıdır, lakin əzələ qlikogeninin parçalanması qandan qlükozanın əzələ emilimini maneə törədir və bununla da digər toxumalarda istifadə üçün mövcud olan qlükoza miqdarını artırır. Qaraciyərdəki qlikogen depoları bədənin hər yerində, xüsusən də mərkəzi sinir sistemində istifadə üçün qlükoza anbarı kimi xidmət edir. İnsan beyni ac və oturaq insanların qanındakı qlükozanın təxminən 60% -ni istehlak edir[4].
Qlikogen, bitkilərdə enerji anbarı kimi fəaliyyət göstərən qlükoza polimeri olan nişastanın analoqudur. Nişastadan daha çox dallı və daha yığcam olan amilopektinə bənzər bir quruluşa malikdir[5]. Hər ikisi quruduqda ağ tozdur. Qlikogen bir çox hüceyrə tipli sitozol sitoplazmada qranullar şəklində olur və qlükoza dövriyyəsində əhəmiyyətli rol oynayır. Qlikogen ani qlükoza tələbatını ödəmək üçün tez bir zamanda səfərbər edilə bilən bir enerji ehtiyatı təmin edir, lakin triqliserid (lipid) enerji mağazalarından daha az yığcamdır. Beləliklə, eyni zamanda bir çox parazitar protozoa üçün bir saxlama ehtiyatıdır[6].
Tərkibi


Qlikogen, orta zəncir uzunluğu təxminən 8–12 qlükoza vahidi və bir qlikogen molekulu başına 2.000–60.000 qalıq olan qlükoza qalıqlarının xətti zəncirlərindən ibarət olan dallı bir biopolimerdir[7].
Qlikogenin sintez üsulu səbəbindən hər bir qlikogen qranulu zülal qlikogeninə əsaslanır[8].
Əzələ, qaraciyər və yağ hüceyrələrindəki qlikogen, qlikogenin hər qramı üçün 0,45 millimol (18 mq) kaliumla bağlanmış üç və ya dörd hissədən ibarət sudan ibarət olan nəmləndirilmiş formada saxlanılır[9].
Qlükoza, yüksək konsentrasiyalarda osmotik təzyiqə dərin təsir göstərə bilən və hüceyrədə heç bir dəyişiklik etmədən saxlanılarsa hüceyrənin zədələnməsinə və ya ölümünə səbəb ola biləcək bir osmotik molekuldur. Qlikogen osmotik olmayan bir molekuldur, buna görə də osmotik təzyiqi pozmadan qlükozanı hüceyrədə saxlamaq üçün bir həll olaraq istifadə edilə bilər[10] .
Funksiyalar
Bədəndə, xüsusən də əzələlərdə və qaraciyərdə saxlanılan qlikogen miqdarı əsasən məşqdən, bazal metabolik sürətdən və qidalanma vərdişlərindən asılıdır (xüsusən də tip 1 oksidləşdirici liflər)[11]. İstirahət edən əzələlərin qlikogen səviyyələri, mövcud hissəciklərin ölçüsünü artırmaq əvəzinə qlikogen hissəciklərinin sayını dəyişdirməklə əldə edilir, baxmayaraq ki, istirahət edən qlikogen hissəciklərinin əksəriyyəti nəzəri maksimumdan azdır. Kiçik miqdarda qlikogen böyrəklər, qırmızı qan hüceyrələri, ağ qan hüceyrələri və beyindəki qlial hüceyrələr də daxil olmaqla digər toxumalarda və hüceyrələrdə olur. Hamiləlik dövründə uterus, embrionu qidalandırmaq üçün qlikogen də saxlayır[12].
Qaraciyər
Karbohidrat və ya protein tərkibli qidalar yeyildikdə və həzm edildikdə, qanda qlükoza səviyyəsi yüksəlir və insulin pankreas tərəfindən ifraz olunur[13][14]. İnsulin, qlikogen sintezi də daxil olmaqla bir neçə fermentin fəaliyyətini stimullaşdıraraq hepatositlərə təsir göstərir. Qlükoza molekulları, insulin və qlükoza çox olduğu müddətdə glikogen zəncirlərinə əlavə olunur. Bu yeməkdən sonra və ya "dolğun" vəziyyətdə qaraciyər qandan çıxardığından daha çox qlükoza alır[15].
Yemək həzm edildikdən və qlükoza səviyyəsi aşağı düşməyə başladıqdan sonra insulin ifrazı azalır və qlikogen sintezi dayanır. Enerjiyə ehtiyac olduqda, qlikogen parçalanır və yenidən qlükoza çevrilir. Qlikogen fosforilaz, qlikogenin parçalanmasında əsas fermentdir. Qarşıdakı 8–12 saat ərzində, qaraciyər qlikogendən əldə edilən qlükoza, qanın əsas qlükoza qaynağıdır və bədənin qalan hissəsinin yanacaq üçün istifadə edir[16].
Pankreas tərəfindən istehsal olunan başqa bir hormon olan qlükagon, bir çox cəhətdən insulinə qarşı siqnal kimi xidmət edir. Normaldan aşağı insulin səviyyəsinə cavab olaraq (qanda qlükoza səviyyəsi normal həddən aşağı düşməyə başlayanda), qlükagon artan miqdarda ifraz olunur və həm glikogenolizi (glikogenin parçalanması), həm də qlükoneogenezi (digər mənbələrdən qlükoza istehsalı) stimullaşdırır[17].
Əzələ
Əzələ hüceyrələri üçün qlikogen, əzələ hüceyrələri üçün mövcud qlükozanın birbaşa ehtiyat mənbəyi kimi çıxış edir. Az miqdarda olan digər hüceyrələr də lokal olaraq istifadə edirlər. Əzələ hüceyrələrində qlükozanın qana keçməsi üçün lazım olan qlükoza-6-fosfataz olmadığından, saxladıqları qlikogen yalnız daxili istifadə üçün mövcuddur və digər hüceyrələrə köçürülmür[18]. Bu, qaraciyər hüceyrələrindən fərqli olaraq, lazım gəldikdə, yığılmış qlikogeni qlükozaya asanlıqla parçalayır və digər orqanlara enerji vermək üçün qandan keçir[19].
Tarixi
Qlikogen Klaude Bernard tərəfindən kəşf edilmişdir[20] . Təcrübələri göstərdi ki, qaraciyərdə "bir fermentin" təsiri ilə şəkərin səviyyəsinin azalmasına səbəb ola biləcək bir maddə var[21]. 1857-ci ilə qədər "la matière qlycogène" və ya "şəkər əmələ gətirən maddə" adlandırdığı bir maddənin salınmasını təsvir etmişdi. Qaraciyərdə qlikogen aşkar edildikdən qısa müddət sonra A.Sanson əzələ toxumasında qlikogen də olduğunu aşkar etdi[22].
Klinik uyğunluq
Qlikogen mübadiləsinin pozulması
Qlikogen mübadiləsinin pozulduğu ən çox görülən vəziyyət, anormal miqdarda insulin səbəbiylə qaraciyərdə qlikogenin yığılması və ya anormal şəkildə tükənə biləcəyi diabetdir. Normal qlükoza metabolizmasının bərpası ümumiyyətlə qlikogen metabolizmasını da normallaşdırır[23].
Həddindən artıq insulinin səbəb olduğu hipoqlikemiyada qaraciyərdə qlikogen səviyyəsi yüksəkdir, lakin yüksək insulin səviyyəsi qan şəkərinin normal səviyyəsini qorumaq üçün lazım olan qlikogenolizi maneə törədir. Qlükaqon bu tip hipoqlikemiya üçün ümumi bir müalicədir[24] .
Müxtəlif anadangəlmə metabolik xəstəliklər, qlikogenin sintezi və ya parçalanması üçün lazım olan fermentlərin olmamasından qaynaqlanır. Bunlara toplu olaraq qlikogen saxlama xəstəlikləri deyilir[25].
Qlikogen tükənməsi və dözümlülük məşqi
Maraton qaçışçıları, xizəkçilər və velosipedçilər kimi uzun məsafəli idmançılar, kifayət qədər karbohidrat qəbul etmədən uzun müddət idman etdikdən sonra demək olar ki, bütün idmançıların qlikogen anbarları tükəndikdə qlikogen tükənməsi ilə qarşılaşırlar[26].
Qlikogen tükənməsinin qarşısını üç yolla almaq olar[27]:
- Birincisi, karbohidratlar məşq zamanı qan qlükozasına (yüksək qlisemik indeks) mümkün olan ən yüksək sürətlə çevrilərək daim istehlak olunur[28].
- İkincisi, dözümlülük və xüsusi rejimləri (məsələn, oruc, aşağı intensivlik dözümlülük) uyğunlaşdıraraq, bədən yanacaq kimi istifadə olunan yağ turşularının faizini artırmaq üçün həm yanacaq səmərəliliyini, həm də məşq qabiliyyətini artırmaq üçün I tip əzələ liflərini şərtləndirə bilər[29][30].
- Üçüncüsü, məşq və ya pəhriz nəticəsində qlikogen anbarlarını tükətdikdən sonra çox miqdarda karbohidratlar istehlak edərək, bədən əzələdaxili qlikogen anbarlarının tutumunu artıra bilər[31]. Bu proses karbohidrat yüklənməsi kimi tanınır. Ümumiyyətlə, müvəqqəti qlikogen tükənməsi nəticəsində əzələ insulinə həssaslıq artdığından karbohidrat mənbəyinin qlisemik indeksi heç bir əhəmiyyət kəsb etmir[32]. İdmançılar gərgin məşqdən sonra karbohidratlar və kofein istehlak etdikdə, onların qlikogen ehtiyatları daha tez dolmağa meyllidir[33].
İstinadlar
- McArdle, William D.; Katch, Frank I.; Katch, Victor L. Exercise physiology: Energy, nutrition, and human performance (6th). Lippincott Williams & Wilkins. 2006. səh. 12. ISBN 978-0-7817-4990-9. 2022-04-07 tarixində arxivləşdirilib. İstifadə tarixi: 2021-09-21.
- Sadava, David E.; Purves, William K.; Hillis, David M.; Orians, Gordon H.; Heller, H. Craig. Life (9th, International). W. H. Freeman. 2011. ISBN 9781429254311.
- Berg JM, Tymoczko JL, Gatto GJ, Stryer L. Biochemistry (Eighth). New York: W. H. Freeman. 2015-04-08. ISBN 9781464126109. OCLC 913469736.
- Wasserman DH. "Four grams of glucose". American Journal of Physiology. Endocrinology and Metabolism. 296 (1). January 2009: E11–21. doi:10.1152/ajpendo.90563.2008. PMC 2636990. PMID 18840763.
Four grams of glucose circulates in the blood of a person weighing 70 kg. This glucose is critical for normal function in many cell types. In accordance with the importance of these 4 g of glucose, a sophisticated control system is in place to maintain blood glucose constant. Our focus has been on the mechanisms by which the flux of glucose from liver to blood and from blood to skeletal muscle is regulated. ... The brain consumes ∼60% of the blood glucose used in the sedentary, fasted person. ... The amount of glucose in the blood is preserved at the expense of glycogen reservoirs (Fig. 2). In postabsorptive humans, there are ∼100 g of glycogen in the liver and ∼400 g of glycogen in muscle. Carbohydrate oxidation by the working muscle can go up by ∼10 fold with exercise, and yet after 1 h, blood glucose is maintained at ∼4 g.
- Guyton, Arthur C.; Hall, John Edward. Guyton and Hall Textbook of Medical Physiology. New York, New York: Saunders/Elsevier. 2011. ISBN 978-5-98657-013-6. 2021-09-13 tarixində arxivləşdirilib. İstifadə tarixi: 2021-09-21.
- Bergström, Jonas; Hermansen, Lars; Hultman, Eric; Saltin, Bengt. "Diet, Muscle Glycogen and Physical Performance". Acta Physiologica Scandinavica (ingilis). 71 (2–3). 1967: 140–150. doi:10.1111/j.1748-1716.1967.tb03720.x. ISSN 1365-201X. PMID 5584523. 2021-08-30 tarixində arxivləşdirilib. İstifadə tarixi: 2021-09-21.
- Jensen, Rasmus; Ørtenblad, Niels; Stausholm, Marie-Louise Holleufer; Skjærbæk, Mette Carina; Larsen, Daniel Nykvist; Hansen, Mette; Holmberg, Hans-Christer; Plomgaard, Peter; Nielsen, Joachim. "Heterogeneity in subcellular muscle glycogen utilisation during exercise impacts endurance capacity in men". The Journal of Physiology (ingilis). 598 (19). 2020: 4271–4292. doi:10.1113/JP280247. ISSN 1469-7793. PMID 32686845 (#bad_pmid). 2022-04-06 tarixində arxivləşdirilib. İstifadə tarixi: 2021-09-21.
- Jensen, Rasmus; Ørtenblad, Niels; Stausholm, Marie‐Louise H.; Skjærbæk, Mette C.; Larsen, Daniel N.; Hansen, Mette; Holmberg, Hans‐Christer; Plomgaard, Peter; Nielsen, Joachim. "Glycogen supercompensation is due to increased number, not size, of glycogen particles in human skeletal muscle". Experimental Physiology (ingilis). 106 (5). May 2021: 1272–1284. doi:10.1113/EP089317. ISSN 0958-0670. PMID 33675088 (#bad_pmid).
- Jensen, Rasmus; Ørtenblad, Niels; Stausholm, Marie‐Louise H.; Skjærbæk, Mette C.; Larsen, Daniel N.; Hansen, Mette; Holmberg, Hans‐Christer; Plomgaard, Peter; Nielsen, Joachim. "Glycogen supercompensation is due to increased number, not size, of glycogen particles in human skeletal muscle". Experimental Physiology (ingilis). 106 (5). May 2021: 1272–1284. doi:10.1113/EP089317. ISSN 0958-0670. PMID 33675088 (#bad_pmid).
- Marchand, I.; Chorneyko, K.; Tarnopolsky, M.; Hamilton, S.; Shearer, J.; Potvin, J.; Graham, T. E. "Quantification of subcellular glycogen in resting human muscle: granule size, number, and location". Journal of Applied Physiology (ingilis). 93 (5). 2002-11-01: 1598–1607. doi:10.1152/japplphysiol.00585.2001. ISSN 8750-7587. PMID 12381743.
- Moses SW, Bashan N, Gutman A. "Glycogen metabolism in the normal red blood cell". Blood. 40 (6). December 1972: 836–843. doi:10.1182/blood.V40.6.836.836. PMID 5083874.
- Ingermann RL, Virgin GL. "Glycogen content and release of glucose from red blood cells of the sipunculan worm themiste dyscrita" (PDF). J Exp Biol. 129. 1987: 141–149. doi:10.1242/jeb.129.1.141.
- Miwa I, Suzuki S. "An improved quantitative assay of glycogen in erythrocytes". Annals of Clinical Biochemistry. 39 (Pt 6). November 2002: 612–13. doi:10.1258/000456302760413432. PMID 12564847.
- Murray, Bob. "Fundamentals of glycogen metabolism for coaches and athletes". Nutrition Reviews. 76 (4). April 2018: 243–259. doi:10.1093/nutrit/nuy001. PMC 6019055. PMID 29444266.
- Oe Y, Baba O, Ashida H, Nakamura KC, Hirase H. "Glycogen distribution in the microwave-fixed mouse brain reveals heterogeneous astrocytic patterns". Glia. 64 (9). June 2016: 1532–1545. doi:10.1002/glia.23020. PMC 5094520. PMID 27353480.
- Campbell, Neil A.; Williamson, Brad; Heyden, Robin J. Biology: Exploring Life. Boston, MA: Pearson Prentice Hall. 2006. ISBN 978-0-13-250882-7. 2014-11-02 tarixində arxivləşdirilib. İstifadə tarixi: 2021-09-21.
- Ryley, J.F. "Studies on the metabolism of the protozoa. 5: Metabolism of the parasitic flagellate Trichomonas foetus". The Biochemical Journal. 59 (3). March 1955: 361–369. doi:10.1042/bj0590361. PMC 1216250. PMID 14363101.
- Benchimol, Marlene; Elias, Cezar Antonio; de Souza, Wanderley. "Tritrichomonas foetus: Ultrastructural localization of calcium in the plasma membrane and in the hydrogenosome". Experimental Parasitology. 54 (3). December 1982: 277–284. doi:10.1016/0014-4894(82)90036-4. ISSN 0014-4894. PMID 7151939.
- Mielewczik, Michael; Mehlhorn, Heinz; al Quraishy, Saleh; Grabensteiner, E.; Hess, M. "Transmission electron microscopic studies of stages of histomonas meleagridis from clonal cultures". Parasitology Research (ingilis). 103 (4). 1 September 2008: 745–750. doi:10.1007/s00436-008-1009-1. ISSN 0932-0113. PMID 18626664.
- Young, F.G. "Claude Bernard and the discovery of glycogen". British Medical Journal. 1 (5033). 22 June 1957: 1431–1437. doi:10.1136/bmj.1.5033.1431. JSTOR 25382898. PMC 1973429. PMID 13436813.
- Manners, David J. "Recent developments in our understanding of glycogen structure". Carbohydrate Polymers. 16 (1). 1991: 37–82. doi:10.1016/0144-8617(91)90071-J. ISSN 0144-8617.
- Ronner, Peter. Netter's Essentials Biochemistry. USA: Elsevier. 2018. 254. ISBN 978-1-929007-63-9.
- Berg, Tymoczko, & Stryer. Biochemistry (7th, International). W. H. Freeman. 2012. səh. 338. ISBN 978-1429203142.
- Berg; və b. Biochemistry (7th, International). W. H. Freeman. 2012. səh. 650.
- Kreitzman SN, Coxon AY, Szaz KF. "Glycogen storage: Illusions of easy weight loss, excessive weight regain, and distortions in estimates of body composition" (PDF). The American Journal of Clinical Nutrition. 56 (1, Suppl). 1992: 292s–293s. doi:10.1093/ajcn/56.1.292S. PMID 1615908. 2015-06-18 tarixində arxivləşdirilib (PDF). İstifadə tarixi: 2021-09-21.
- "Glycogen Biosynthesis; Glycogen Breakdown". oregonstate.edu. 2021-05-12 tarixində arxivləşdirilib. İstifadə tarixi: 2018-02-28.
- Stryer, L. Biochemistry (3rd). Freeman. 1988. səh. 451.
- "Glycogen Biosynthesis; Glycogen Breakdown". oregonstate.edu. 2021-05-12 tarixində arxivləşdirilib. İstifadə tarixi: 2018-02-28.
- "Methods of endurance training, Part 1". 2009-10-30. 2018-07-22 tarixində orijinalından arxivləşdirilib. İstifadə tarixi: 2021-09-21.
- "Steady state vs. tempo training and fat loss". 2008-06-02. 2017-09-05 tarixində orijinalından arxivləşdirilib. İstifadə tarixi: 2021-09-21.
- McDonald, Lyle. "Research review: An in-depth look into carbing up on the cyclical ketogenic diet". 2012-07-25. 2020-11-11 tarixində arxivləşdirilib. İstifadə tarixi: 19 February 2017.
- McDonald, Lyle. The Ketogenic Diet: A complete guide for the dieter and the practitioner. Lyle McDonald. 1998.
- Beelen, M.; Burke, L.M.; Gibala, M.J.; van Loon, L.J.C. "Nutritional strategies to promote post-exercise recovery". International Journal of Sport Nutrition and Exercise Metabolism. 20 (6). December 2010: 515–532. doi:10.1123/ijsnem.20.6.515. PMID 21116024.