Lipopolisaxarid
Lipopolisaxarid (ing. Lipopolysaccharide; rus. Липополисахарид)- Lipoqlikan və endotoksin kimi tanınan lipololisaxaridlər (LPS) lipidA və O-antigen, kovalent rabitəli xarici və daxili nüvədən ibarət böyük molekullardır; onlar Gram-mənfi bakteriyaların xarici membranda aşkar edilir və heyvanlarda güclü immun sistemindən xəbər verir. Lipoliqosaxarid (LOS) termini bakterial lipopolisaxaridlərin aşağı molekulyar çəki formasını adlandırmaq üçün istifadə olunur.

Kəşf
LPS zəhərli fəaliyyəti ilk dəfə Riçard Friderik Johannes Pfeiffer tərəfindən aşkar olundu və o exotoksinlərdən fərqli olaraq “endotoksin” adlandırıldı, o qeyd etdi ki, toksinlər bakteriya məhv olan zaman bakteriyalar tərəfindən ətraf mühitə yayılır, endotoxinlər isə bakterial hüceyrə "çərçivəsində" saxlanılan toksinlərdir və yalnız bakterial hüceyrə divarının dağıdılmasından sonra ətrafa yayıla bilir.[1]
Sonrakı tədqiqatlar göstərdi ki, qram mənfi bakteriyalar olan LPS-in ətrafa yayılması bakterial hüceyrə divarının dağıdılması zəruriliyini tələb etmir, əksinə, LPS başqa zəhərlilik faktorlarını və zülalları özündə saxlayan bilən bakterial xarici membran qovuqcuqlarının (OMVs) formasında iştirak edən membran qovuqcuğunun normal fizioloji fəaliyyətinin hissəsi kimi gizlədilir.[2]
Son zamanlar “endotoksin” termini LPS-in sinonimi kimi istifadə olunur, buna baxmayaraq LPS-lə uyğun olmayan digər bakteriyalarla ifraz olunan bir neçə molekullar vardır ki, onlar Bacillus thuringiensislə ifraz olunan delta-endotoksin zülallarıdır.[3]
Bakteriyada funksiyaları
LPS bakteriyaların struktur bütövlüyünün köməyi və membranı müxtəlif kimyəvi hücumlardan qoruyan gram-mənfi bakteriyaların xarici membranının əsas komponentidir. LPS eyni zamanda membran divarının mənfi yükünü artırır və ümumi membran strukturunu sabitləşdirməyə kömək edir. O, qram - mənfi bakteriya üçün mühüm əhəmiyyət kəsb edir, hansı ki məhv olduqda ya şəklini, ya da yerini dəyişir.
LPS bir əksendotoksindir və normal heyvan immun sistemindən kəskin fərqlənir. O, həmçinin bakterial ekologiyanın yerüstü yapışma, baktereoloji həssaslıq və amöb kimi yırtıcılar ilə qarşılıqlı əlaqədə olan qeyri-patogen aspektlər ilə qarışdırılır. LPS Omptin fəaliyyətinin düzgün uyğunlaşdırılması üçün tələb olunur; lakin, sərbəst LPS omptinlərə mane olacaq.
Tərkibi
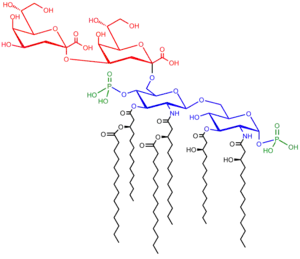
Bu üç hissədən ibarətdir:
- . O antigen (və ya O polisaxarid)
- . Nüvə oliqosaxarid
- . Lipid A
O – antigen
Bir təkrar qlikan polimer LPS –lərə istiqamətlənmiş bakteriyaların O antigen, O polisaxarid və ya O yan-zəncirdən ibarətdir. O antigen nüvə oliqosaxaridə hücum edir və LPS molekulunun xarici mühitini əhatə edir. O zəncirin tərkibi gərginlikdən gərginliyə doğru dəyişir. Məsələn, E. koli növləri tərəfindən istehsal edilən 160-dən çox müxtəlif O antigen strukturları vardır.[4] O zəncirinin varlığı və ya olmaması LPS kobud və ya hamar hesab olunub-olunmadığını müəyyən edir. O-zəncirlər ümumi uzunluğu LPS-in hamarlığını göstərəcək, halbuki O-zəncirlərin olmaması və ya azalması LPS nahamar göstərəcək.[5] Nahamar LPS-li bakteriyaların adətən membrane divarından hidrofobik antibiotiklərə keçidi vardır ki, elə bu vaxtdan nahamar LPS daha hidrofobikdirlər.[6] O antigen bakterial hüceyrənin xarici səthində təhlükəyə məruz qalır və məntiqi nəticə olaraq antikor kütləsi ilə tanınması üçün bir hədəf deyil.
Nüvə
Nüvə mühiti həmişə lipid A-ya birbaşa hücum edən oliqosaxarid komponentindən və əsasən heptose və 3-deoxy-D-mannooctulosonic turşusu kimi şəkərdən (həmçinin KDO, keto-deoxyoctulosonate kimi tanınan[7]) ibarətdir. Bir çox bakteriya ların LPS nüvəsi həmçinin fosfat, amin turşuları, və etanolamin substansiyaları kimi qeyri-karbohidrogen komponentlərindən ibarətdir.
Lipid A
Lipid A normal şəraitdə yağ turşuları ilə əhatələnmiş fosforlu qlukosamin disaxariddir. Bu hidrofob yağ turşusu zəncirləri LPS bakterial membrana daxil edir və LPS layihələri hüceyrə səthinə əsaslanır. Lipid A mühiti qram-mənfi bakteriyaların toksikliyinə görə məsuliyyət daşıyır. Bakterial hüceyrələr immun sistemində olan zaman, membran olan lipid A fraqmentləri od, ishal və mümkün məhvedici endotoksik şokdan (həmçinin septik şok) azad olur. Lipid A parçası LPS-in mühafizə komponentidir.[8]
Lipooliqosaxaridlər
Lipooliqosaxaridlər (LOS) belə Neisseria spp və Haemophilus spp kimi gram mənfi bakteriyaların bəzi növlərinin xarici membranında aşkar olunan qlikolipidlərdir. Termin bakterial LPS aşağı molekulyar çəki forması ilə sinonimidir.[9] LOS qram mənfi zərflərin xarici membran bütövlüyü və funksionallığının mərkəzi rolunu oynayır. LOS müəyyən bakterial infeksiyalar patogenezinin mühüm rolunu oynayır, cünki onlar immunostimulatorlar və immunomodulatorlar kimi hərəkət edə bilir.
Bundan əlavə, LOS molekulları bəzi bakterial növlərin immun müdafiə qabiliyyəti üçün yardım, molekulyar mimika və antigenik müxtəlifliyi keçirmək üçün məsuliyyət daşıyırlar və beləliklə bu bakterial növlərin xəstəlik törətməsinə köməklik edirlər. Kimyəvi olaraq lipooliqosaxaridlər O-antigenlərin çatışmazlığı və yalnız bir lipid A-özüllü xarici membran-lövbər parça və oliqosaxarid əsasdır.[10] Neisseria meningitidis halında lipid A molekulunun bir hissəsi simmetrik quruluşa malikdir və daxili nüvə 3-deoxy-D-Manno-2-octulosonic turşusu (KDO) və heptose (Hep) siniflərindən ibarətdir. Xarici nüvə oligqosaxarid zəncir bakterial gərginlikdən asılı olaraq dəyişir.[10]
Lipooliqosaxarid termini bakterial lipopolisaxaridlərin aşağı molekulyar çəki şəklində istinad üçün istifadə olunan iki forması kimi təsnif edilə bilər: yüksək molekulyar çəki (Mr və ya kobud) forması yüksək molekulyar çəkiyə malikdir, O-zəncir poliaxaridini təkrarlayır, aşağı molekuyar çəki forması zamanı O-zəncirinin çatışmamazlığı bu yerdə qısa oliqosaxaridə malik olur.[9]
LPS dəyişikliklər
LPS-in yaradılması xüsusi şəkər strukturunu təqdim etmək üçün dəyişdirilə bilər. O həmçinin digər bir LPS və ya qlikosiltranferlər (LPS toksinlerin mane imkan verir) tərəfindən qəbul edilə bilər, hansi ki onlar müxtəlif şəkərləri adlandırmaq üçün o şəkər strukturları kimi istifadə oluna bilər. Bu yaxınlarda nümayiş etdirildi ki, bağırsaqdakı xüsusi ferment (qələvi fosfataza) LPS-i LPS karbohidratlarında tapılan iki fosfat qruplarının aradan qaldırılması ilə zərərsizləşdirir.[11] Bu kütlə qram - mənfi bakteriyaların adətən kiçik bağırsaqda aşkar olunan potensial zəhərli təsirini idarə etmək üçün bir adaptiv mexanizm kimi fəaliyyət göstərə bilər.[12]
Biosintez və daşıma
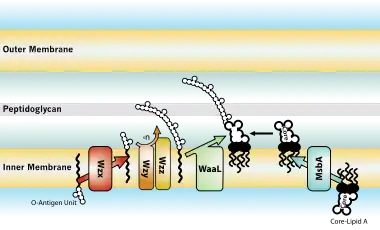
Qram-mənfi bakteriyalarla yoluxma zamanı bioloji təsirlər
İmmun cavab
LPS-in hərəkəti prototip endotoksin kimi CD14/TLR4/MD2 reseptor kompleksini bir çox hüceyrə növləridə bağlayır, ancaq xüsusilə monositlər, dentritik hüceyrələr, makrofaqlar və B hüceyrələr hansı ki, onlar pro-iltihab sitokinlərin, azot oksidi və eikosanoidlərin ifraz olunmasına köməklik göstərir.[15] LPS-lər eyni zamanda ekzogen pirogendir (xarici oyanma-induksiya maddələri). Mühüm əhəmiyyət kəsb edən qram-mənfi bakteriyalar, bu molekulları yeni antimikrob agentləri üçün hədəflərə çevirir. Bəzi tədqiqatçılar bütün lipopolisaxaridlərin sianobakteriya üçün əsas zəhərli təsiri barədə ümumiləşdirilmiş hesabat vermişlər.[16]
LPS funksiyası bir çox transkripsiya amillərinin fəaliyyətində eksperimental tədqiqat rolunu oynamışdır. LPS həmçinin septik şokda vasitəçilərin bir çox növlərini istehsal edir. İnsanlar LPS qarşı digər heyvanlara nisbətən daha çox həssasdır (məsələn, siçan). İnsanlarda şokun dozası 1 µg/kg, lakin siçan min dəfə çox dozaya qədər dözə bilir.[17] Bu iki növ arasında təbii antikor dövriyyə səviyyəsində fərqlər ola bilər.[18][19] Deyilənlər göstərir ki, LPS CD4 T- hüceyrələrinin yayılmasının IL-10 – tabeli dayanmasına səbəb olur və monositlərdə tənzimləyici PD-1 səviyyəsi funksiyası PD-1-dən PD-L-ə qədər monositlər tərəfindən bağlanmadan sonra IL-10 məhsulu üstünlük təşkil edir.
Endotoksinlər patogen qram-mənfi bakteriyalı infeksiyaların dramatik klinik təzahürləri üçün böyük hissədir, belə infeksiyalara Neisseria meningitidis, meningococcemia, Waterhouse-Frideriksen sindromu və meningit aiddir. Bruce Beutler TLR4-ün LPS reseptor olduğunu sübut etməklə 2011-ci ildə Psixologiya və Tibb sahəsində Nobel mükafatına sahib oldu.[20]
LPS-dən olan bir neçə bakterial növlər insanın hüceyrə səthli molekulların kütləsinə kimyəvi oxşar olmasını göstərir; bəzi bakteriyaların qabiliyyəti öz səthlərində molekulları təqdim edir, hansı ki, kimyəvi bənzər və ya eyni bəzi hüceyrə kütlələrinin molekul səthlərinə molekulyar bənzərliklə şərtlənir.[21] Məsələn, Neisseria meningitidis L 2, 3, 5, 7, 9-da oliqosaxaridin son tetrasaxarid hissəsi (lacto-N-neotetraose) paraqlobosiddə aşkar olunan eyni tetrasaxariddir, ABH qlikolipid antigenləri üçün xəbərçi insan eritrositlərində aşkar olunur.[9]
Başqa misal olaraq, patogen Neisseria spp olan oligosaxaridin son trisaxarid hissəsidir (lactotriaose). LOS həmçinin insan hüceyrələrinin laktoneoseriyalarının qlikoşipinqolipidlərində tapılır. B və C qruplarının bir çox meninqokokiləri qonokokkidən çox yaxşıdır, onlar bu trisaxaridin öz LOS strukturunun bir hissəsi kimi göstərirlər. Bu insan hüceyrəsinin olması bundan başqa immun sistemindən bir “kamuflyaj” kimi hərəkət edərək, müəyyən insan leykosit antigen (HLA) HLA-B35 kimi genotipləri ilə zəhərləndiyində immun davamlılığının ləğvində rol oynaya bilər.
İmmun cavabının dəyişkənlik təsiri
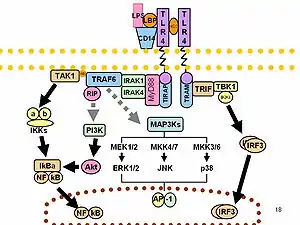
O-antigenlər (xarici karbohidratlar) LPS molekulunun antigenik xüsusiyyətə malik ən dəyişkən hissəsdir. Əksinə, lipid A ən güclü mühafizə hissəsidir. Lakin, lipid A-nın tərkibi dəyişkəndir. Bu dəyişikliklərin bəziləri bu LPS-in antaqonist xassələrini yaya bilər. Məsələn Rodobakter sphaeroides diphosphoryl lipid A-sı (RsDPLA) insan hüceyrələrində LPS-in güclü rəqibidir, lakin dağsiçanı və atların hüceyrələrində bir mübarizə vasitəsidir. Fərziyyə olunur ki, konusvari Lipid A (məsələn, E. koli) çox agonistikdirlər, daha az Porfiromonas gingivalis kimi konusvari Lipid A müxtəlif siqnallar verə bilər (TLR2 əvəzinə TLR4) və Rodobakter sphaeroides kimi silindrik lipid A TLRs-ə qarşı antaqonistdir.[22][23]
LPS gen qrupları müxtəlif növlərin, yarımnövlərin, bitki və heyvanların bakterial pathogen növləri arasında çox dəyişkəndir.[24][25] Normal insan qan mayesi anti-LOS bakterisiddən ibarətdir ki, onlar bakteriosid və dözümlüdür, onların infeksiyaya səbəb olan növləri ilə müqayisədə normal maye öz spesifikliyi ilə fərqlənən anti-LOS-a malik olurlar.[26]
Bu fərqlər əsasən LOS molekulunun oliqosaxarid hissəsinin tərkibində LOS molekul quruluşuna aid edilə bilər. Neisseria gonorrhoeae-də LOS molekulların antigenic səbəbiylə qabiliyyəti infeksiya zamanı dəyişə bilər ki, bu bakteriyalar LOS bir çox növü üçün faza dəyişkənliyi kimi tanınan xarakterik sintez kimi təqdim olunur.[26] Bundan əlavə, Neisseria gonorrhoeae, habelə Neisseria meningitidis və Haemophilus influenza vitroda öz LOS şəklini dəyişir, məsələn sialilation olmadan (sialik turşusu qalıqları ilə modifikasiyası) və nəticədə onların müqavimətini artırmaq üçün öldürülmək və ya fəaliyyətini zəiflətmək və ya bakterisid antikor təsirdən yayınmaq lazımdır.[9]
Sialilation həmçinin immunitet sistemi hüceyrələrinə, habelə aşağı oksidləşdirici partlayışa mane olan neytrofillərə əlavə və faqositoz kömək edə bilər. Mal-qarada rast gəlinən patogen Haemophilus somnus həmçinin LOS mərhələsi dəyişikliyini göstərir, xarakterik olaraq iribuynuzluların immun müdafiəsində yayınmadan kömək edə bilər. Bu müşahidələr göstərir ki, bakterial yerüstü molekulların dəyişiklikləri LOS patogen kimi humoral (antikor və vasitəçiliyi tamamlayır) və hüceyrə- vasitəçiliyi (məsələn, neytrofillərin tərəfindən məhv edilməsi) ilə immun müdafiəyə kömək edə bilər.[26]
İstinadlar
- Parija SC. Textbook of Microbiology & Immunology. India: Elsevier. Jan 1, 2009. ISBN 8131221636. December 3, 2016 tarixində arxivləşdirilib. İstifadə tarixi: April 4, 2015.
- Kulp A, Kuehn MJ. "Biological functions and biogenesis of secreted bacterial outer membrane vesicles". Annu. Rev. Microbiol. 64. 2010: 163–84. doi:10.1146/annurev.micro.091208.073413. PMC 3525469. PMID 20825345.
- Rietschel ET, Kirikae T, Schade FU, Mamat U, Schmidt G, Loppnow H, və b. "Bacterial endotoxin: molecular relationships of structure to activity and function". FASEB J. 8 (2). 1994: 217–25. PMID 8119492.
- Raetz CR, Whitfield C. "Lipopolysaccharide endotoxins". Annu. Rev. Biochem. 71. 2002: 635–700. doi:10.1146/annurev.biochem.71.110601.135414. PMC 2569852. PMID 12045108.
- Rittig MG, Kaufmann A, Robins A, Shaw B, Sprenger H, Gemsa D, və b. "Smooth and rough lipopolysaccharide phenotypes of Brucella induce different intracellular trafficking and cytokine/chemokine release in human monocytes". J. Leukoc. Biol. 74 (6). 2003: 1045–55. doi:10.1189/jlb.0103015. PMID 12960272.
- Tsujimoto H, Gotoh N, Nishino T. "Diffusion of macrolide antibiotics through the outer membrane of Moraxella catarrhalis". J. Infect. Chemother. 5 (4). 1999: 196–200. doi:10.1007/s101569900025. PMID 11810516.
- Hershberger C, Binkley SB. "Chemistry and metabolism of 3-deoxy-D-mannooctulosonic acid. I. Stereochemical determination". J. Biol. Chem. 243 (7). 1968: 1578–84. PMID 4296687.
- Tzeng YL, Datta A, Kolli VK, Carlson RW, Stephens DS. "Endotoxin of Neisseria meningitidis composed only of intact lipid A: inactivation of the meningococcal 3-deoxy-D-manno-octulosonic acid transferase". J. Bacteriol. 184 (9). May 2002: 2379–88. doi:10.1128/JB.184.9.2379-2388.2002. PMC 134985. PMID 11948150.
- Moran AP, Prendergast MM, Appelmelk BJ. "Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease". FEMS Immunol. Med. Microbiol. 16 (2). 1996: 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.
- Kilár A, Dörnyei Á, Kocsis B. "Structural characterization of bacterial lipopolysaccharides with mass spectrometry and on- and off-line separation techniques". Mass Spectrom Rev. 32 (2). 2013: 90–117. doi:10.1002/mas.21352. PMID 23165926.
- Bates JM, Akerlund J, Mittge E, Guillemin K. "Intestinal alkaline phosphatase detoxifies lipopolysaccharide and prevents inflammation in zebrafish in response to the gut microbiota". Cell Host Microbe. 2 (6). 2007: 371–82. doi:10.1016/j.chom.2007.10.010. PMC 2730374. PMID 18078689.
- Munford R, Lu M, Varley AW. "Kill the Bacteria...and Also Their Messengers?". Advances in Immunology. 103. 2009: 29. doi:10.1016/S0065-2776(09)03112-8. PMID 19755182. (#accessdate_missing_url)
- Wang X, Quinn PJ. "Lipopolysaccharide: Biosynthetic pathway and structure modification". Prog. Lipid Res. 49 (2). 2010: 97–107. doi:10.1016/j.plipres.2009.06.002. PMID 19815028.
- Ruiz N, Kahne D, Silhavy TJ. "Transport of lipopolysaccharide across the cell envelope: the long road of discovery". Nat. Rev. Microbiol. 7 (9). 2009: 677–83. doi:10.1038/nrmicro2184. PMC 2790178. PMID 19633680.
- Abbas, Abul. Basic Immunology. Elsevier. 2006. ISBN 978-1-4160-2974-8.
- Stewart I, Schluter PJ, Shaw GR. "Cyanobacterial lipopolysaccharides and human health - a review". Environ Health. 5. 2006: 7. doi:10.1186/1476-069X-5-7. PMC 1489932. PMID 16563160.
- Warren HS, Fitting C, Hoff E, Adib-Conquy M, Beasley-Topliffe L, Tesini B, və b. "Resilience to bacterial infection: difference between species could be due to proteins in serum". J. Infect. Dis. 201 (2). 2010: 223–32. doi:10.1086/649557. PMC 2798011. PMID 20001600.
- Reid RR, Prodeus AP, Khan W, Hsu T, Rosen FS, Carroll MC. "Endotoxin shock in antibody-deficient mice: unraveling the role of natural antibody and complement in the clearance of lipopolysaccharide". J. Immunol. 159 (2). 1997: 970–5. PMID 9218618.
- Boes M, Prodeus AP, Schmidt T, Carroll MC, Chen J. "A critical role of natural immunoglobulin M in immediate defense against systemic bacterial infection". J. Exp. Med. 188 (12). 1998: 2381–6. doi:10.1084/jem.188.12.2381. PMC 2212438. PMID 9858525.
- Said EA, Dupuy FP, Trautmann L, Zhang Y, Shi Y, El-Far M, və b. "Programmed death-1-induced interleukin-10 production by monocytes impairs CD4+ T cell activation during HIV infection". Nat. Med. 16 (4). 2010: 452–9. doi:10.1038/nm.2106. PMC 4229134. PMID 20208540.
- Chastain EM, Miller SD. "Molecular mimicry as an inducing trigger for CNS autoimmune demyelinating disease". Immunol. Rev. 245 (1). 2012: 227–38. doi:10.1111/j.1600-065X.2011.01076.x. PMC 3586283. PMID 22168423.
- Netea MG, van Deuren M, Kullberg BJ, Cavaillon JM, Van der Meer JW. "Does the shape of lipid A determine the interaction of LPS with Toll-like receptors?". Trends Immunol. 23 (3). 2002: 135–9. doi:10.1016/S1471-4906(01)02169-X. PMID 11864841.
- Seydel U, Oikawa M, Fukase K, Kusumoto S, Brandenburg K. "Intrinsic conformation of lipid A is responsible for agonistic and antagonistic activity". Eur. J. Biochem. 267 (10). 2000: 3032–9. doi:10.1046/j.1432-1033.2000.01326.x. PMID 10806403.
- Reeves PP, Wang L. "Genomic organization of LPS-specific loci". Curr. Top. Microbiol. Immunol. Current Topics in Microbiology and Immunology. 264 (1). 2002: 109–35. doi:10.1007/978-3-642-56031-6_7. ISBN 978-3-540-42682-0. PMID 12014174.
- Patil PB, Sonti RV. "Variation suggestive of horizontal gene transfer at a lipopolysaccharide (lps) biosynthetic locus in Xanthomonas oryzae pv. oryzae, the bacterial leaf blight pathogen of rice". BMC Microbiol. 4. 2004: 40. doi:10.1186/1471-2180-4-40. PMC 524487. PMID 15473911.
- Yamasaki R, Kerwood DE, Schneider H, Quinn KP, Griffiss JM, Mandrell RE. "The structure of lipooligosaccharide produced by Neisseria gonorrhoeae, strain 15253, isolated from a patient with disseminated infection. Evidence for a new glycosylation pathway of the gonococcal lipooligosaccharide". J. Biol. Chem. 269 (48). 1994: 30345–51. PMID 7982947.
Xarici keçidlər
- Lipopolisaxarid mövzusu ilə bağlı Tibbi subyekt rubrikası.