

Un potencial d'aición, tamién llamáu impulsu llétricu, ye una onda de descarga llétrica que viaxa a lo llargo de la membrana celular modificando la so distribución de carga llétrica.[1] Los potenciales d'aición utilizar nel cuerpu pa llevar información ente unos texíos y otros, lo que fai que sían una carauterística microscópica esencial pa la vida. Pueden xenerase por diversu tipos de célules corporales, pero les más actives nel so usu son les célules del sistema nerviosu pa unviar mensaxes ente célules nervioses (sinapsis) o dende célules nervioses a otros texíos corporales, como'l músculu o les glándules.
Munches plantes tamién xeneren potenciales d'aición que viaxen al traviés del floema pa coordinar la so actividá. La principal diferencia ente los potenciales d'aición d'animales y plantes ye que les plantes utilicen fluxos de potasiu y calciu ente que los animales utilicen potasiu y sodiu.
Los potenciales d'aición son la vía fundamental de tresmisión de códigos neurales. Les sos propiedaes pueden frenar el tamañu de cuerpos en desenvolvimientu y dexar el control y coordinación centralizaos d'órganos y texíos.
Considerancies xenerales
Siempres hai una diferencia de potencial o un potencial de membrana ente la parte interna y la parte esterna de la membrana celular (polo xeneral, de 70 mV). ~La carga d'una membrana celular inactiva caltener en valores negativos (l'interior respectu al esterior) y varia dientro d'unos estrechos márxenes. Cuando'l potencial de membrana d'una célula escitable se despolariza más allá d'un ciertu estragal (de –65 mV a –55 mV app) la célula xenera (o dispara) un potencial d'aición. Ye importante esclariar que tanto l'interior como l'esterior celular caltiénense electroneutros, esto ye, nun hai una diferencia de carga neta ente l'interior de la célula y l'esterior. La diferencia de potencial de membrana deber a la distribución diferencial d'iones (mayoritariamente, cloru y sodiu nel esterior celular, y potasiu y aniones orgánicos nel interior).
| Concentraciones ióniques trasmembrana d'una célula mamífera[2] | |||
|---|---|---|---|
| Tipu | Ion | Concentración esterna (mM) | Concentración interna (mM) |
| Catiónica | Sodiu (Na+) | 145 | 5-15 |
| Potasiu (K+) | 5 | 140 | |
| Magnesiu (Mg2+) | 1-2 | 0,5 | |
| Calciu (Ca2+) | 1-2 | 0,0001 | |
| Hidróxenu (H+) | 0,00004 | 0,00007 | |
| Aniónica | Cloru (Cl-) | 110 | 5-15 |
Bien básicamente, un potencial d'aición ye un cambéu bien rápido na polaridá de la membrana de negativu a positivu y vuelta a negativu, nun ciclu que dura unos milisegundos. Cada ciclu entiende una fase ascendente, una fase descendente y d'últimes una fase hiperpolarizada. Nes célules especializaes del corazón, como les célules del marcapasos coronariu, la fase pandu de voltaxe entemediu puede apaecer antes de la fase descendente.
Los potenciales d'aición midir con téuniques de rexistru d'electrofisioloxía (y más apocayá, con neurochips de MOSFET). Un osciloscopiu que rexistre'l potencial de membrana d'un puntu concretu d'un axón amuesa cada etapa del potencial d'aición, ascendente, descendente y refractaria, a midida que la onda pasa. Estes fases xuntes formen un arcu sinusoidal deformado. La so amplitú depende d'ónde algamó'l potencial d'aición al puntu de midida y el tiempu trescurríu.
El potencial d'aición nun se caltién nun puntu de la membrana plasmática, sinón que viaxa a lo llargo de la membrana. Puede movese a lo llargo d'un axón a muncha distancia, por casu tresportando señales dende'l celebru hasta l'estremu de la migollu espinal. N'animales grandes como les xirafes o les ballenes la distancia pue ser de dellos metros.
La velocidá, frecuencia y simplicidá de los potenciales d'aición varia según el tipu celular ya inclusive ente célules del mesmu tipu. Aun así, los cambeos de voltaxe tienden a tener la mesma amplitú ente elles. Nuna mesma célula, dellos potenciales d'aición consecutivos son práuticamente indistinguibles. La "causa" del potencial d'aición ye l'intercambiu d'iones al traviés de la membrana celular. Primero, un estímulu abre les canales de sodiu. Yá que hai dellos iones de sodiu nel esterior, y l'interior de la neurona ye negativu con rellación al esterior, los iones de sodiu entren rápido a la neurona. El sodiu tien una carga positiva, asina que la neurona vuélvese más positiva y empieza a despolarizarse. Les canales de potasiu retrásense un pocu más n'abrir; una vegada abiertos el potasiu sale rápido de la célula, revirtiendo la despolarización. Más o menos nesti momentu, les canales de sodiu empiecen a cerrase, llogrando que'l potencial d'aición vuelva a –70 mV (repolarización). En realidá'l potencial d'aición va más allá de –70 mV (hiperpolarización), por cuenta de que les canales de potasiu quédense abiertos un pocu más. Gradualmente les concentraciones d'iones tornen a los niveles de reposu y la célula vuelve a –70 mV.
Mecanismu subxacente

Potencial de membrana en reposu
Cuando la célula nun ta aguiyada por corrientes despolarizantes supraumbrales, dizse que s'atopa nun potencial de membrana en reposu.
La membrana celular ta compuesta mayoritariamente por una bicapa de fosfolípidos altamente hidrofóbica, que torga'l pasu llibre de partícules cargaes como los iones. Polo cual esta bicapa de fosfolipidos pórtase como un condensador, dixebrando cargues (daes polos iones en disolución) a una distancia d'aproximao 4 nm. Esto dexa la mantención del potencial de membrana a lo llargo del tiempu. El potencial de membrana deber a la distribución diferencial d'iones ente l'interior y l'esterior celular. Esti potencial de membrana ye calteníu a lo llargo del tiempu pol tresporte activu d'iones per parte de bombes, tales como la bomba sodio-potasiu y la bomba de calciu. Estes proteínes usen la enerxía d'hidrólisis d'ATP pa tresportar iones en contra del so gradiente electroquímicu, calteniendo asina los gradientes de concentraciones ióniques que definen el potencial de membrana.
Fases del potencial d'aición
Les variaciones del potencial de membrana mientres el potencial d'aición son resultáu de cambeos na permeabilidá de la membrana celular a iones específicos (en concretu, sodiu y potasiu) y por consiguiente cambeos nes concentraciones ióniques nos compartimientos intracelular y estracelular. Estes rellaciones tán matemáticamente definíes pola ecuación de Goldman, Hodgkin y Katz (GHK).
![{\displaystyle Y_{m,K_{x}Na_{1-x}Cl}={\frac {RT}{F}}\ln {\left({\frac {P_{Na^{+}}[Na^{+}]_{\mathrm {out} }+P_{K^{+}}[K^{+}]_{\mathrm {out} }+P_{Cl^{-}}[Cl^{-}]_{\mathrm {in} }}{P_{Na^{+}}[Na^{+}]_{\mathrm {in} }+P_{K^{+}}[K^{+}]_{\mathrm {in} }+P_{Cl^{-}}[Cl^{-}]_{\mathrm {out} }}}\right)}}](../I/07fb5829676adca451c900d4fb47b785eaa93078.svg)
Los cambeos na permeabilidá de la membrana y l'establecimientu y cese de corrientes ióniques mientres el potencial d'aición reflexen l'apertura y zarru de les canales ióniques que formen zones de camín al traviés de membrana pa los iones. Les proteínes que regulen el pasu d'iones al traviés de la membrana respuenden a los cambeos de potencial de membrana.
Nun modelu simplificáu del potencial d'aición, el potencial de reposu d'una parte de la membrana caltener cola canal de potasiu. La fase ascendente o de despolarización del potencial d'aición empecípiase cuando la canal de sodiu dependiente de potencial ábrese, faciendo que la permeabilidá del sodiu supere llargamente a la del potasiu. El potencial de membrana va escontra ENa. En delles célules, como les célules del marcapasos coronariu, la fase ascendente xenerar por concentración de calciu más que de sodiu.
Tres un curtiu intervalu, la canal de potasiu dependiente de voltaxe (tardiegu) ábrese, y la canal de sodiu ciérrase gradualmente. De resultes, el potencial de membrana vuelve al estáu de reposu, amosáu nel potencial d'aición como una fase descendente.
Por cuenta de que hai más canales de potasiu abiertes que canales de sodiu (les canales de potasiu de membrana y canales de potasiu dependiente de voltaxe tán abiertes, y la canal de sodiu ta zarrada), la permeabilidá al potasiu ye agora enforma mayor qu'antes del entamu de la fase ascendente, cuando solo les canales de potasiu de membrana taben abiertes. El potencial de membrana averar a EK más de lo que taba en reposu, faciendo que'l potencial tea en fase refractaria. La canal de potasiu tardiego dependiente de voltaxe ciérrase por cuenta de la hiperpolarización, y la célula torna a la so potencial de reposu.
Estragal y iniciación

Los potenciales d'aición desencadénense cuando una despolarización inicial algama un estragal. Esti potencial estragal varia, pero de normal ta en redol a –55 a –50 milivoltios sobre'l potencial de reposu de la célula, lo qu'implica que la corriente d'entrada d'iones sodio supera la corriente de salida d'iones potasio. El fluxu neto de carga positiva qu'acompaña los iones sodiu despolariza el potencial de membrana, desaguando nuna apertura de los canal de sodiu dependientes de voltaxe. Estes canales apurren un fluxu mayor de corrientes ióniques escontra l'interior, aumentando la despolarización nuna retroalimentación positiva que fai que la membrana llegue a niveles de despolarización elevaos.
L'estragal del potencial d'aición puede variar camudando l'equilibriu ente les corrientes de sodiu y potasiu. Por casu, si dalgunos de los canal de sodiu tán inactivos, determináu nivel de despolarización va abrir menos canales de sodiu, y aumenta asina l'estragal de despolarización necesariu pa empecipiar el potencial d'aición. Este ye'l principiu del funcionamientu del periodu refractariu (ver periodu refractariu).
Los potenciales d'aición son bien dependientes de los equilibrios ente iones sodio y potasiu (anque hai otros iones que contribúin minoritariamente a los potenciales, como calciu y cloru), y por ello los modelos fáense utilizando solu dos canales ióniques transmembrana: una canal de sodiu dependiente de voltaxe y una canal de potasiu pasivo. L'orixe del estragal del potencial d'aición puede visualizase na curva I/V (imaxe) que representa les corrientes ióniques al traviés de les canales frente al potencial de membrana. (La curva I/V representada na imaxe ye una rellación instantánea ente corrientes. Amuésase'l picu de corrientes a determináu voltaxe, rexistráu primero qu'asoceda nenguna inactivación (1 ms n'algamando esi voltaxe pal sodiu). Tamién ye importante apuntar que la mayoría de voltaxes positivos del gráficu solo pueden consiguise per medios artificiales, por aciu l'aplicación d'electrodos a les membranes).
Destaquen cuatro puntos na curva I/V indicaos poles fleches de la figura:
- La flecha verde indica'l potencial de reposu de la célula y el valor del potencial d'equilibriu pal potasiu (Ek). Por cuenta de que la canal K+ ye l'únicu abiertu con esos valores de voltaxe negativos, la célula va caltener en Ek. Va Apaecer un potencial de reposu estable con cualquier voltaxe en que'l sumatorio I/V (llinia verde) encruz el puntu de corriente nula (exa x) con una pendiente positiva, como fai na flecha verde. Esto ye por cuenta de que cualquier perturbación del potencial de membrana escontra valores negativos va significar corrientes netes d'entrada que despolarizarán la célula más allá del puntu d'encruz, ente que cualquier perturbación escontra valores positivos va significar corrientes netes de salida que hiperpolarizarán la célula. Asina, cualquier cambéu del potencial de membrana de rimada positiva tiende a devolver a la célula al valor d'encruz cola exa.
- La flecha mariella indica l'equilibriu del potencial Na+ (ENa). Nesti sistema de dos iones, el ENa ye la llende natural del potencial de membrana del que la célula nun puede pasar. Los valores de corrientes nel gráficu qu'entepasen esta llende hanse Unidá de midida midíu de forma artificial obligando a la célula a devasalo. Aun así, el ENa solo podría algamar si la corriente de potasiu nun esistiera.
- La flecha azul indica'l voltaxe máximu que puede algamar el picu del potencial d'aición. Ye'l potencial de membrana máximu que la célula n'estáu natural puede algamar, y nun puede llegar al ENa por cuenta de la aición contraria de los fluxos de potasiu.
- La flecha colorada indica l'estragal del potencial d'aición. Ye'l puntu onde l'Isum camudar a un fluxu neto escontra l'interior. Destaca que nesti puntu traviésase'l puntu de fluxu neto cero, pero con rimada negativa. Cualesquier "puntu de corte con rimada negativa" del nivel de fluxu cero nel gráficu I/V ye un puntu inestable. Si'l voltaxe nesti puntu ye negativu, el fluxu va escontra l'esterior y la célula tiende a volver al potencial de reposu. Si'l voltaxe ye positivu, el fluxu va escontra l'interior y tiende a despolarizar la célula. Esta despolarización implica mayor fluxu escontra l'interior, faciendo que los fluxos de sodiu se realimenten. El puntu nel que la llinia verde algama'l valor más negativu ye cuando toles canales de sodiu tán abiertos. La despolarización más allá d'esti puntu baxa les corrientes de sodiu una y bones la fuercia llétrico mengua a midida que el potencial de membrana averar a ENa.
L'estragal del potencial d'aición confundir dacuando col estragal de l'apertura de canales de sodiu. Ye una incorreición, una y bones les canales de sodiu escarecen d'estragal. Otra manera, ábrense en respuesta a la despolarización aleatoriamente. La despolarización nun implica tantu l'apertura de les canales como la medría de la probabilidá de que s'abrir. Inclusive en potenciales de hiperpolarización, una canal de sodiu puede abrir esporádicamente. Amás, l'estragal del potencial d'aición nun ye'l voltaxe a la que'l fluxu d'iones sodio faise importante; ye'l puntu en qu'entepasa'l fluxu de potasiu.
Biolóxicamente, nes neurones la despolarización aniciar nes sinapsis dendríticas. En principiu, los potenciales d'aición podríen xenerase en cualquier puntu a lo llargo de la fibra nerviosa. Cuando Luigi Galvani afayó la eletricidá animal faciendo que la pierna d'una xaronca muerta volviera a la vida tocando'l nerviu ciáticu con un escalpelu, aplicándo-y ensin dase cuenta una carga electrostática negativa y empecipiando un potencial d'aición.
Modelu del circuitu
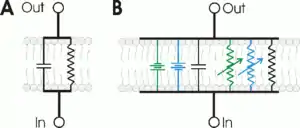
Les membranes celulares con canales ióniques pueden representase con un modelu de circuitu RC pa entender meyor l'espardimientu de potenciales d'aición en membranes biolóxiques. Nestos circuitos, la resistencia representa les canales ióniques de membrana, ente que el condensador representa l'aislamientu de la membrana lipídica. Los potenciómetros indiquen les canales ióniques regulaos por voltaxe, yá que'l so valor camuda col voltaxe. Una resistencia de valor fixu representa les canales de potasiu que caltienen el potencial de reposu. Los gradientes de sodiu y potasiu indicar nel modelu como fontes de voltaxe (pila).
Espardimientu
Nos axones amielínicos, los potenciales d'aición arrobínense como una interaición pasiva ente la despolarización que se mueve pola membrana y les canales de sodiu regulaos por voltaxe.

Cuando una parte de la membrana celular se despolariza lo suficiente como por que se abrir les canales de sodiu dependientes de voltaxe, los iones de sodiu entren na célula por espardimientu facilitáu. Una vegada dientro, los iones positivos de sodiu impulsen los iones próximos a lo llargo del axón por repulsión electrostática, y atraen los iones negativos dende la membrana axacente.
Como resultancia, una corriente positiva mover a lo llargo del axón, ensin que nengún ion tea moviéndose bien rápido. Una vegada que la membrana axacente ta abondo despolarizada, les sos canales de sodiu dependientes de voltaxe ábrense, realimentando el ciclu. El procesu repitir a lo llargo del axón, xenerándose un nuevu potencial d'aición en cada segmentu de la membrana.
Velocidá d'espardimientu
Los potenciales d'aición arrobínense más rápidu en axones de mayor diámetru, si los demás parámetros caltiénense. La principal razón por qu'asoceda ye que la resistencia axial de la lluz del axón ye menor cuanto mayor sía'l diámetru, por cuenta de la mayor rellación ente superficie total y superficie de membrana nuna corte tresversal. Como la superficie de la membrana ye la torga principal pal espardimientu del potencial en axones amielínicos, la medría d'esta tasa ye una forma especialmente efectiva d'amontar la velocidá de la tresmisión.
Un exemplu d'un animal qu'utiliza l'aumentu de diámetru de axón como regulador de la velocidá d'espardimientu del potencial de membrana ye'l calamar xigante. El axón del calamar xigante controla la contraición muscular acomuñada cola respuesta de fuximientu de depredadores del animal. Esti axón puede devasar 1 mm de diámetru, y posiblemente sía una adautación pa dexar una activación bien rápida del mecanismu d'escape. La velocidá de los impulsos nerviosos nestes fibres ye una de les más rápides de la naturaleza, pa los que tienen neurones amielínicas.
Conducción saltatoria
En axones mielínicos, la conducción saltatoria ye'l procesu pol que los potenciales d'aición paecen saltar a lo llargo del axón, siendo refechos solo nunos aniellos ensin aisllar (los nodos de Ranvier). La conducción saltatoria amonta la velocidá de conducción nerviosa ensin tener qu'amontar significativamente el diámetru del axón nel sistema nerviosu.
Desempeñó un papel importante na evolución d'organismos más complexos que los sos sistemes nerviosos precisen tresmitir rápido potenciales d'aición a llargues distancies. Ensin conducción saltatoria, la velocidá de conducción riquiría medríes drástiques nel diámetru del axón, a tal puntu que podríen resultar na formación de sistemes nerviosos descomanadamente grandes pa los cuerpos que tienen d'agospialos.
Mecanismu detalláu
La principal torga pa la velocidá de tresmisión en axones amielínicos ye la capacitancia de la membrana. La capacidá d'un condensador puede menguase baxando'l área d'una corte tresversal de les sos plaques, o amontando la distancia ente les plaques. El sistema nerviosu utiliza la mielinización p'amenorgar la capacitancia de la membrana. La mielina ye una vaina proteutora creada alredor de los axones poles célules de Schwann y los oligodendrocitos, célules de la neuroglía qu'entartallen el so cuerpu celular y el so axón respeutivamente formando llámines de membrana y plasma. Estes llámines arollar nel axón, alloñando les plaques conductores (el plasma intra y estracelular) ente sigo, menguando la capacitancia de la membrana.
L'aislamientu resultante redunda nuna conducción rápida (práuticamente instantánea) de los iones al traviés de les seiciones mielinizadas del axón, pero torga la xeneración de potenciales d'aición nestos segmentos. Los potenciales d'aición solo vuélvense a producir nos nodos de Ranvier desmielinizados, que s'asitien ente los segmentos mielinizados. Nestos aniellos hai un gran númberu de canales de sodiu dependientes de voltaxe (hasta cuatro órdenes de magnitú cimera a la densidá de axones amielínicos), que dexen que los potenciales d'aición refacer de forma eficaz nellos.
Por cuenta de la mielinización, los segmentos aisllaos del axón actúen como un cable pasivu: conducen los potenciales d'aición rápido porque la capacitancia de la membrana ye bien baxa, y embriven la degradación de los potenciales d'aición porque la resistencia de la membrana ye alta. Cuando esta señal que s'arrobina de forma pasiva algama un nodo de Ranvier, empecipia un potencial d'aición que viaxa de nuevu de forma pasiva hasta qu'algama'l siguiente nodo, repitiendo'l ciclu.
Minimización de daños
El llargor de los segmentos mielinizados d'un axón ye importante pa la conducción saltatoria. Tienen De ser tan llargos como sía posible pa optimizar la distancia de la conducción pasiva, pero non lo suficiente como por que el amenorgamientu na intensidá de la señal sía tanta que non algame l'estragal de sensibilidá nel siguiente nodo de Ranvier. En realidá, los segmentos mielinizados son lo suficientemente llargos por que la señal que s'arrobina pasivamente percuerra siquier dos segmentos calteniendo una amplitú de señal abonda como pa empecipiar un potencial d'aición nel segundu o tercer nodo. Asina s'alza'l factor de seguridá de la conducción saltatoria, dexando que la tresmisión trespase nodos en casu de que tean estropiaos.
Enfermedá
Dalgunes enfermedaes afecten la conducción saltatoria y mengüen la velocidá de desplazamientu d'un potencial d'aición. La más conocida de toes estes enfermedaes ye la esclerosis múltiple, na que los daños na mielina imposibilitan el movimientu coordináu.
Periodu refractariu
Defínese como'l momentu nel que la célula escitable nun respuende ante un estímulu y polo tanto nun xenera un nuevu potencial d'aición. Estremar en dos: periodu refractariu absolutu (o efectivu) y periodu refractariu relativu.
El periodu refractariu absolutu ye aquel nel que les canales de Na+ sensibles a voltaxe atópense inactivos, polo que se inhibe el tresporte d'iones sodio. Sicasí, el periodu refractariu relativu dase ayundes de la fase de repolarización, onde les canales de Na+ pasu ente pasu empiecen a reactivase. D'esta manera, al amestar un estímulu excitatorio bien intensu puede provocase que les canales (que s'atopen zarraos nesi momentu) ábranse y xeneren un nuevu potencial d'aición que la so amplitú depende de cuántu s'avere nesi momentu'l potencial de membrana al potencial de reposu.[3] El periodu refractariu relativu termina dempués de la fase de hiperpolarización (o postpotencial) onde toles canales de Na+ sensibles a voltaxe tán zarraos y disponibles pa un nuevu estímulu.
Tamién esiste un periodu refractariu efectivu, que solo se repara nes célules musculares cardiaques (esto debe a que les célules atópense formando un sincitio celular). Nesti casu, la célula se despolariza de normal, pero nun puede conducir dichu estímulu a les célules vecines a ella. Esti periodu refractariu ye un parámetru bien útil na evaluación de drogues antiarrítmicas.
El periodu refractariu varia de célula a célula, y ye una de les carauterístiques que dexen dicir si una célula ye más o menos escitable qu'otra. N'otros casos como'l músculu cardiacu, el so ampliu periodu refractariu déxa-y la capacidá de non tetanizarse.
Ventaya evolutiva
Con al respective de les causes poles que la naturaleza desenvolvió esta forma de comunicación, esto respuéndese al considerar la tresmisión d'información a gran distancia al traviés d'un axón nerviosu. Pa llevar la información d'un estremu a otru del axón, han de superase lleis físiques como les que condicionen el movimientu de señales llétriques nun cable. Por cuenta de la resistencia llétrica y a la capacitancia del cable, les señales tienden a degradase a lo llargo del cable. Estes propiedaes denominaes propiedaes del cable determinen les llendes físiques a los que pueden llegar les señales.
El correutu funcionamientu del cuerpu precisa que les señales lleguen d'un estremu a otru del axón ensin perdes nel camín. Un potencial d'aición non solo arrobínase a lo llargo del axón, sinón que se refai pol potencial de membrana y corrientes ióniques en cada estrechamientu de la membrana nel so camín. N'otres pallabres, la membrana nerviosa refai'l potencial de membrana en tola so amplitú a midida que la señal percuerre'l axón, superando les llendes qu'impon la teoría de llinies de tresmisión.
Ver tamién
- Potencial de membrana
- Excitabilidad neuronal
- Hiperpolarización
- Potencial de membrana
- Potencial excitatorio postsináptico
- Sinapsis
Referencies
- ↑ Berg, Jeremy Mark; Stryer, Lubert; Tymoczko, John L. (2007). potencial+de+acci%C3%B3n+impulso+el%C3%A9ctrico&hl=es&sa=X&ved=0ahUKEwjZ1M637anXAhXIShQKHSM1BQQQ6AEIOjAD#v=onepage&q=Un%20potencial%20de%20acci%C3%B3n%20impulsu%20el%C3%A9ctrico&f=false Bioquímica (en castellanu). Reverte. ISBN 9788429176001. Consultáu'l 6 de payares de 2017.
- ↑ Alberts, Bruce. Bioloxía Molecular de la Célula, páx. 542.
- ↑ Cingolani, A. E.; Houssay, A. B. (2000) Fisioloxía Humana. L'Atenéu, páx. 250. ISBN 978-950-02-0376-0.
Bibliografía
Fuentes xenerales
- Bear, M. F., Connors, B. W., Paradiso, M.A: Neurociencia: esplorando'l celebru. Barcelona: Masson, 2002. ISBN 84-458-1259-9
- Kandel, E. R., Schwartz, J. H., Jessell, T. M.: Principios de neurociencia. Madrid: McGraw-Hill, 2001, 4ª ed. ISBN 84-486-0311-7
- Purves. D., et al: «Ion Channels Underlying Action Potentials.» En Neuroscience. Sunderland, Massachusetts: Sinauer Associates, 2004, tercer ed. Disponible en llinia
Fontes primaries
- Clay, J. R. «Axonal excitability revisited.» Prog Biophys Mol Biol. 2005 May;88(1):59-90. PMID 15561301
- Hodgkin, A. L. y A. F. Huxley. «Currents carried by sodium and potassium ions through the membrane of the giant axon of Loligo.» J Physiol. 1952 Apr;116(4):449-72. PMID 14946713
- Hodgkin, A. L. y A. F. Huxley. «The components of membrane conductance in the giant axon of Loligo.» J Physiol. 1952 Apr;116(4):473-96. PMID 14946714
- Hodgkin, A. L. y A. F. Huxley. «The dual effect of membrane potential on sodium conductance in the giant axon of Loligo.» J Physiol. 1952 Apr;116(4):497-506. PMID 14946715
- Hodgkin, A. L. y A. F. Huxley. «A quantitative description of membrane current and its application to conduction and excitation in nerve.» J Physiol. 1952 Aug;117(4):500-44. PMID 12991237
Enllaces esternos
- «El potencial d'aición: hestoria del so descubrimientu y estudiu.» Por Fidel Ramón y Jesús Hernández-Falcón. Universidá Nacional Autónoma de Méxicu: Facultá de medicina.
N'inglés
- Demostración de fluxu iónico mientres el potencial d'aición.
- Simulador d'ecuaciones Nernst/Goldman.
- La ecuación de Nerst y los potenciales d'aición nel sistema nerviosu.
| Control d'autoridaes |
|
|---|
 Datos: Q194277
Datos: Q194277 Multimedia: Action potentials
Multimedia: Action potentials