| Flavonoide | |
|---|---|
 | |
| Xeneral | |
| Propiedaes físiques | |
| Propiedaes termoquímiques | |
| Peligrosidá | |
| Identificadores | |
Flavonoide (del llatín flavus, "mariellu") ye'l términu xenéricu con que s'identifica a una serie de metabolitos secundarios de les plantes. Son sintetizaos a partir d'una molécula de fenilalanina y 3 de malonil-CoA, al traviés de lo que se conoz como "vía biosintética de los flavonoides", que'l so productu, la estructura base, se cicla gracies a una enzima isomerasa. La estructura base, una cadarma C6-C3-C6, puede sufrir darréu munchos cambeos y amiestes de grupos funcionales, polo que los flavonoides son una familia bien diversa de compuestos, anque tolos productos finales carauterizar por ser polifenólicos y solubles n'agua. Los flavonoides que caltienen la so cadarma pueden clasificase, según les isomerizaciones y los grupos funcionales que-yos son amestaos, en 6 clases principales: les chalcones, les flavones, los flavonoles, los flavandioles, les antocianines, y los taninos entestaos.[1] más una séptima clase, les aurones, teníes en cuenta por dellos autores por tar presentes nuna cantidá considerable de plantes. Tamién la cadarma puede sufrir cambeos, convirtiéndose entós na cadarma de los isoflavonoides o'l de los neoflavonoides, que polo tanto tamién son derivaos de los flavonoides.
Los flavonoides se biosintetizan en toles "plantes terrestres" o embriofites, y tamién en delles algues Charophyta, y anque toles especies comparten la vía biosintética central, tienen una gran variabilidá na composición química de los sos productos finales y nos mecanismos de regulación de la so biosíntesis, polo que la composición y concentración de flavonoides ye bien variable ente especies y en respuesta al ambiente. Los flavonoides son sintetizaos nel citoplasma y depués migren escontra'l so destín final nes vacuoles celulares. Cumplen funciones metabóliques importantes nes plantes, delles funciones son comunes a toles plantes y otres son específiques de dalgunos taxones. Como exemplu de funciones universales, los flavonoides son responsables de la resistencia de les plantes a la fotooxidación de la lluz ultravioleta del Sol, intervienen nel tresporte de la hormona auxina, y créese que funcionen como defensa ante'l herbivorismo. Una función importante cumplida en munches plantes ye l'atraición de los animales polinizadores, al traviés del color o'l golor que dan a la planta o a les sos flores.
Los flavonoides adquirieron notoriedá pública arriendes de la so actividá biolóxica nel home, que los consume colos vexetales. Los flavonoides tienen propiedaes bien apreciaes en medicina, como antimicrobianos, anticancerígenos, amenorgamientu del riesgu d'enfermedaes cardiaques, ente otros efeutos. Tamién son conocíos polos cultivadores de plantes ornamentales, que manipolien l'ambiente de les plantes p'aumentar la concentración de flavonoides que dan el color a les fueyes y a les flores.
Por cuenta de les importantes funciones metabóliques que los flavonoides tienen nes plantes y los animales, les sos víes biosintéticas y mecanismos de regulación tán siendo curioso estudiaos. La ciencia aplicada aprovechó esta conocencia en munchos trabayos d'inxeniería metabólica, nos que se buscó por casu, aumentar la concentración de flavonoides beneficiosos nes plantes de consumu humanu o d'usu farmacéuticu, modificar la so concentración en flores ornamentales pa camuda-yos el color, y tornar la so producción nel polen pa llograr la esterilidá de los híbridos d'interés comercial. No que fai a la so producción, desenvolvióse con ésitu un cultivu de bacteries que sintetiza flavonoides d'interés humanu.
Los científicos dieron usos variaos a los flavonoides: los xenes de la biosíntesis de flavonoides fueron usaos como ferramienta p'analizar los cambeos nel ADN, son exemplos conocíos el descubrimientu de les lleis de Mendel (que pudo rastrexar l'heriedu de los xenes de los flavonoides que dan el color a los arbeyos), y el descubrimientu de los xenes saltarinos de Barbara McClintock (que al "saltar" escontra un xen d'un flavonoide lo inutilizan y nun s'espresa'l color nel granu de maíz). La estraición ya identificación de flavonoides tamién foi bien usada polos botánicos sistemáticos pa establecer parentescos ente especies de plantes.
Entá queda enforma por investigar de los flavonoides, del so valor melecinal, y del so impautu na nutrición y la salú humana y de los animales. Tamién ye necesariu siguir la investigación de la so estructura, el so metabolismu y el so biodisponibilidad, polo que s'esperen importantes progresos nesti campu[2]
 La estructura base de los flavonoides tien la cadarma d'una chalcona, y l'aición de la enzima isomerasa convertir nuna flavanona. |
 Cadarma de los isoflavonoides..  Cadarma de los neoflavonoides. |
Descubrimientu
Probablemente la primer vegada que la ciencia describió a los flavonoides foi cuando Robert Boyle en 1664 fixo una primer descripción de los efeutos de los pigmentos de les flores en mediu ácidu y en mediu básicu.[1]
El primer flavonoide foi identificáu en 1930 pol premiu Nobel de Fisioloxía y Medicina Szent-Györgyi, quien aislló del pulgu de llimón una sustancia, la citrina, que probó regular la permeabilidá de los capilares al ser consumida.
Estructura química
El primer flavonoide sintetizáu pola "vía biosintética de los flavonoides" ye una chalcona, que'l so cadarma ye un aniellu bencénico xuníu a una cadena propánica que ta xunida de la mesma a otru aniellu bencénico. Na mayoría de los flavonoides, la cadena de reaiciones sigue, polo que la cadena carbonada que xune los aniellos arumosos se cicla por aición d'una enzima isomerasa, creando una flavanona.
Munches vegaes la biosíntesis sigue y los productos finales, tamién flavonoides, queden xuníos a bien diversos grupos químicos, por casu los flavonoides glucosidados porten molécules d'azucres o los sos derivaos. Tamién pueden encontarse flavonoides parcialmente polimerizados dando llugar a dímeros, trímeros, o complexos multienlazados, como los taninos entestaos.
Tolos flavonoides tienen les carauterístiques de ser polifenólicos y solubles n'agua. Tienen un máximu d'absorción de lluz a los 280 nm.
Estraición y analís
La estraición de los flavonoides realizar con solventes orgánicos d'alta polaridá como'l etanol (el acetato de etilo usar con bastante procuru yá que por calentamientu puede reaccionar con ciertos compuestos). Darréu, realícense estraiciones socesives con solventes de polaridá creciente como: Hexano / Cloroformu (pa flavonoides de baxa polaridá). Acetato de etilo (pa flavonoides de mediana polaridá). Butanol (pa flavonoides d'alta polaridá y, polo xeneral, los de mayor usu farmacolóxicu gracies a la presencia de grupos funcionales de tipu hidroxilo y carbonilo)
La espectrofotometría ye útil p'analizar la concentración de flavonoides nuna sustancia.
Munches vegaes esa midida realízase acoplada a una separación cromatográfica como por casu HPLC.
Clasificación de los flavonoides
Acordies cola nomenclatura de la Unión Internacional de Química Pura y Aplicada.[3] pueden clasificase, según la so cadarma y vía metabólica, en:
- Flavonoides, derivaos de la estructura 2-fenilcromen-4-ona (2-fenil-1,4-benzopirona).
- Isoflavonoides, derivaos de la estructura 3-fenilcromen-4-ona (3-fenil-1,4-benzopirona).
- Neoflavonoides, derivaos de la estructura 4-fenilcumarina (4-fenil-1,2-benzopirona).
 Estructura química de la 2-fenilcromen-4-ona (2-fenil-1,4-benzopirona), cadarma de los flavonoides. |
Estructura química de la 3-fenilcromen-4-ona (3-fenil-1,4-benzopirona), cadarma de los isoflavonoides.. |
Estructura química de la 4-fenilcumarina (4-fenil-1,2-benzopirona), cadarma de los Neoflavonoides. |
Los isoflavonoides formar por migración d'un aniellu bencénico de la posición 2 a 3 del aniellu central. El grupu integra más de 230 estructures, y los dos más conocíos son la genisteína y la daidzeina.[4] La so función ye defender a les plantes del ataque de patóxenos.
Dientro de los flavonoides, reconócense 6 y quiciabes 7 clases principales, según los grupos funcionales que tengan: les chalcones, les flavones, los flavonoles, los flavandioles, les antocianines, los taninos entestaos, y dellos autores consideren tamién a les aurones, qu'otros integren nes chalconas. Tamién hai otros derivaos de los flavonoides que tienen cambeos tales que nun entren dientro de nenguna d'estes clases principales.
El númberu de flavonoides distintos que ye, en teoría, posible ye astronómicu, si tener en cuenta que diez de los carbonos de la cadarma del flavonoide pueden ser sustituyíos por una variedá de grupos distintos, que de la mesma pueden ser hidroxilados, metoxilados, metilados, isoprenilados o benzilados. Amás, cada grupu hidroxilo y dalgunos de los carbonos pueden ser sustituyíos por unu o más azucres distintos, y de la mesma, cada unu d'esos azucres puede ser acilado con una variedá d'acedos fenólicos o alipáticos distintos. S'identificaron y aisllaron alredor de 9.000 flavonoides, pero ensin dulda entá hai munchos más por afayar.[5]
- 1. Les chalconas tán implicaes na estimulación de la polinización gracies a qu'inducen al desenvolvimientu de colores nel espectru de lo visible y nel UV qu'atraen a inseutos (caparines y abeyes)

- 2. Les flavonas son marielles y pueden tar en delles flores, como na prímula, dándo-y un color mariellu a los sos pétalos, o en frutos, como na piel de les uves, yá que son les responsables del color amarellentáu de los vinos blancos. Hai trés flavonas importantes: la tricetina, presente nel polen de delles mirtáceas y tamién nes podocarpáceas (Podocarpus spp.); l'apigenina, presente en munches plantes como la manzanilla, (Matricaria recutita) o'l espín blancu (Crataegus laevigata), da un color marrón marfileño a les flores si preséntase sola; y la luteolina, de color mariellu, qu'inclusive sirve pa tiñir llana y otros texíos, pa lo cual emplegóse la Retama de los tintoreros (Genista tinctoria).
- 3. Los flavonoles suelen ser incoloros o mariellos y atópense nes fueyes y en munches flores. Los más importantes son trés: la quercetina, ye'l flavonol mariellu del polen de munches fagácees (Quercus sp.);[6] la miricetina, presente na uva; y el kaempferol, presente nes inflorescencies y protexer de la lluz ultravioleta. La fisetina ye un flavonol que s'estrayi de la planta del xéneru Amphipterygium.
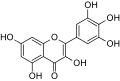


- 4. Hai trés flavandioles carauterísticos: la leucocianidina, presente en delles plantes, como nel plátanu o nel arfueyu criollu (Ligaria cuneifolia); la leucopelargonidina, presente como tal en cierta concentración na alfalfa de secanu (Medicago truncatula); y la leucodelfinidina, que ye activa nel castañu d'indies (Aesculus hippocastanum).
- 5. Les antocianinas, son los pigmentos hidrosolubles presentes nel líquidu vacuolar de les célules responsables de la mayoría de les coloraciones coloraes, azules y violetes de les flores y fueyes.[7]
- 6. Los taninos entestaos son macromolécules constituyíes por unidaes de flavonoides llamaes antocianidinas. Los taninos tán bien llargamente distribuyíos nes plantes como nel té, onde contribúin al sabor astringente.[8]
- 7. Les auronas son responsables de la coloración de delles plantes. A pesar de que se suxurió qu'estos compuestos tán rellacionaos estrechamente coles chalconas, hai pocos nicios alrodiu de les sos víes biosintéticas[9]
- 8. Les flavanonas son precursores d'otros flavonoides más complexos, pero atópense como tales n'altes concentraciones nos cítricos. Les más importantes son: la naringenina, presente nel zusmiu de naranxa, llimón o pomelu, dándo-y un sabor amargoso; la liquiritigenina, presente nel regaliza; y el eriodictiol, presentar nel arbeyu actuando como quimioatrayente pa interactuar con agrobacteries.
- 9. Los dihidroflavonoles son los precursores direutos de flavandioles y flavonoles, pero tamién tienen cierta actividá como tales en delles plantes. Hai trés importantes: la dihidromiricetina, presente nes partes aérees de los uzes (Erica spp.), la dihidroquercetina, nes uves blanques o na alzaparrilla (Smilax aristolochiaefolia); y el dihidrokampferol que puede atopase na madera del pinu siberianu (Pinus sibirica).
Los flavonoides en llantar
Orixe evolutivu
Los flavonoides apaecieron per primer vegada nos ancestros de les embriofitas, qu'entiende al grupu monofilético de toles plantes terrestres (mofos, felechos, gimnospermas y anxospermes). Créese que fueron una de les adautaciones clave pa la transición a la vida terrestre dende l'alga verde ancestral, por cuenta de la so capacidá d'absorber la radiación ultravioleta, muncho más intensa na atmósfera que na agua.[10]
Les enzimes de la biosíntesis de los flavonoides aparentemente derivaron d'enzimes del metabolismu primariu de les plantes, que teníen xenes duplicaos, lo que dexaría l'adautación de dalgunes eses enzimes a otres funciones específiques (Iida et al. 1999[11] Rausher et al. 1999,[12] Durbin et al. 2000.[13]).
La vía biosintética de los flavonoides caltúvose descomanadamente nel intre de la evolución de les plantes, pero hubo considerable diverxencia tantu nos roles que fueron cumpliendo los sos productos finales, como nos mecanismos que regulen la so espresión.[14]
Biosíntesis
.svg.png.webp)
La vía del ácidu shikímico empecipiar nos plastos por condensación de dos productos fotosintéticos, la eritrosa 4-P col fosfoenolpiruvato (PEP), y por diversu cambeos llógrase l'ácidu shikímico, del cual deriven direutamente dellos fenoles nos vexetales. Pero la vía del ácidu shikímico de normal prosigue, y la incorporación d'una segunda molécula de PEP conduz a la formación de fenilalanina.
La vía biosintética de los flavonoides empieza cuando la fenilalanina, por aición de la enzima fenilalanina amonioliasa (PAL) tresformar en acedu cinámico, que depués ye tresformáu n'acedu p-cumarínico por incorporación d'un grupu hidroxilo a nivel d'aniellu arumosu, y l'aición d'una CoA ligasa tresformar en cumaril-SCoA, el precursor de la mayoría de los fenoles d'orixe vexetal, ente los que s'atopen los flavonoides[8]
Regulación de la biosíntesis
- La vía del ácidu shikímico ye dependiente de la lluz.
- L'aición de la fenilalanina amonioliasa, qu'empecipia la vía biosintética de los flavonoides, ye fundamental pa la vida de les plantes y por ello ta puramente regulada. Ente otros factores, la fenilalanina amonioliasa ye activada pola lluz, y depende amás de la concentración de distintes hormones vexetales. L'actividá de la fenilalanina amonioliasa suel aumentar cuando a los vexetales sométese-yos a situaciones d'estrés, como pue ser la falta d'agua ("estrés hídricu"), infeiciones fúngicas o bacterianes, radiaciones UV, y el fríu (por esto últimu les plantes sometíes a baxes temperatures suelen presentar coloraciones acolorataes en tarmos y fueyes, y cuando los iviernos son bien fríos, les flores desenvuelven colores bien intensos na primavera siguiente).
- Hai isozimes dedicaes a la producción de flavonoides distintes en respuesta a señales ambientales distintes.[15]
Funciones nes plantes
- Proteición ante la lluz UV. Los flavonoides incoloros suelen atropase nes capes más superficiales de les plantes y capten hasta'l 90% de les radiaciones UV, torgando los efeutos nocivos d'estes radiaciones nos texíos internos.
- Defensa ante'l herbivorismo. Dellos flavonoides como los taninos, protexen a les plantes xenerando sabores desagradables pa los herbívoros, principalmente amargosos, o testures que pueden resultar desagradables pa los herbívoros, que se ven aguiyaos a escoyer otres plantes.[8]
- Regulación del tresporte de la hormona auxina. Les plantes mutantes que nun tener la enzima chalcona sintasa, que forma parte de la vía biosintética de los flavonoides, amuesen una crecedera irregular por cuenta de un defectu nel tresporte d'auxina al traviés de la planta. Probablemente esi defectu deber a l'ausencia d'esi flavonoide na planta mutante[16]

- Atraición d'animales polinizadores. Munchos flavonoides son componentes de pigmentos de les flores y fueyes que confieren coloraciones atrayentes d'inseutos, colo que la función de munchos flavonoides sería la d'atraer a los polinizadores escontra les flores Un casu bien destacáu, ye'l de les bromeliáceas ente les que s'atopen les especies Tillandsia y Billbergia, que desenvuelven les sos flores sobre un tarmu que se elonga sobre una base con fueyes en roseta. El tarmu elongado ta formáu por una serie de bráctees que presenten un color acoloratáu bien fuerte antes de la polinización o mientres ésta, y depués fáense más verdoses.[17]

- Atraición de preses. Les plantes carnívores, como la Drosera y la Dionaea, tienen antocianinas nes sos flores y fueyes, que cumplen una función d'atraición de los inseutos que los sirven d'alimentu.
- Atraición d'animales dispersores de granes y frutos. Dellos flavonoides confieren arumes y colores a los frutos que los faen más apetecedores pa los herbívoros que s'alimenten d'ellos, cumpliendo asina una función de dispersión de les granes.
- Inducción de la nodulación per parte de les bacteries fijadoras de nitróxenu. Reparóse que'l eriodictiol y la apigenina-7-O-glucósido exudados pol raigañu del arbeyu (Pisum sativum) inducen la nodulación de la agrobacteria Rhizobium leguminosarum. Viose tamién que dos isoflavonoides atopaes en exudados de soya, la daidzeina y la genisteína son inductores de los xenes de la nodulación de delles cepes de Bradyrhizobium japonicum.
- Proteición contra los fungos. La pisatina ye un isoflavonoide producíu pol arbeyu en respuesta a la infeición por fungu ya induz la espresión del xen PDA1 nestos. La pisatina y l'eriodictiol inducen la guañada de les espores de ciertos fungos.
 Estructura molecular de la genisteína.
Estructura molecular de la genisteína.

Aplicaciones de los flavonoides
Aplicaciones en medicina
- Pa un review n'inglés ver Graf et al. 2005.[18]
Los flavonoides consumíos pol home protéxen-y del dañu de los oxidantes, como los rayos UV (que la so cantidá aumenta pel branu); la polución ambiental (minerales tóxicos como'l plomu y el mercuriu); delles sustancies químiques presentes nos alimentos (colorantes, conservantes, etc). Como l'organismu humanu nun tien la capacidá de sintetizar estes sustancies químiques, llograr dafechu de los alimentos qu'inxer.
Nun son consideraos vitamines.
Al llindar l'aición de los radicales llibres (que son oxidantes), los flavonoides amenorguen el riesgu de cáncer, ameyoren los síntomes alérxicos y d'artritis, aumenten l'actividá de la vitamina C, , bloquien la progresión de los tabayones y la dexeneración macular, eviten les tufaradas de calor na menopausia (chornes) y combaten otros síntomes.
Polo xeneral el sabor ye amargosu, llegando inclusive a provocar sensaciones d'astringencia si la concentración de taninos entestaos ye bien alta. El sabor puede variar dependiendo de les sustituciones presentaes na cadarma llegando inclusive a usase como edulcorantes cientos de vegaes más duces que la glucosa.
Los sos efeutos nos humanos pueden clasificase en:
- Propiedaes anticancerosas: munchos demostraron ser tremendamente eficaces nel tratamientu del cáncer. Sábese que munchos inhiben la crecedera de les célules canceroses. Probóse contra'l cáncer de fégadu[19]
- Propiedaes cardiotónicas: tienen un efeutu tónicu sobre'l corazón, potenciando'l músculu cardiacu y ameyorando la circulación. Atribuyíes fundamentalmente al flavonoide quercetina anque apaez en menor intensidá n'otros como la genisteína y la luteolina. Los flavonoides mengüen el riesgu d'enfermedaes cardiaques.[20][21]
- Fraxilidá capilar: ameyoren la resistencia de los capilares y favorecen el qu'éstos non se ruempan, polo que resulten fayadizos pa prevenir el sangráu. Los flavonoides con meyores resultaos nesti campu son la hesperidina, la rutina y la quercetina.
- Propiedaes antitrombóticas: la capacidá d'estos componentes pa torgar la formación de trombos nos vasos sanguíneos fai posible una meyor circulación y una prevención de munches enfermedaes cardiovasculares.
- Amenorgamientu del colesterol: tienen la capacidá de menguar la concentración de colesterol y de triglicéridos.
- Proteición del fégadu: dellos flavonoides demostraron menguar la probabilidá d'enfermedaes nel fégadu. Foi probáu en llaboratoriu que la silimarina protexe y refai el fégadu mientres la hepatitis. Xunto cola apigenina y la quercetina, son bien útiles pa esaniciar ciertes dolencies dixestives rellacionaes col fégadu, como la sensación de plenitú o les vultures.
- Proteición del estómagu: ciertos flavonoides, como la quercetina, la rutina y el kaempferol, tienen propiedaes antiulcéricas al protexer la mucosa gástrica.
- Antiinflamatorios y analxésicos: la hesperidina poles sos propiedaes antiinflamatorias y analxésiques, utilizóse pal tratamientu de ciertes enfermedaes como l'artritis. Los taníns tienen propiedaes astringentes, vasoconstrictoras y antiinflamatorias, pudiéndose utilizar nel tratamientu de les hemorroides.[8]
- Antimicrobianos: isoflavonoides, furanocumarinas y estilbenos demostraron tener propiedaes antibacterianas, antivirales y antifúngicas.
- Propiedaes antioxidantes: Nes plantes los flavonoides actúen como antioxidantes, especialmente les catequinas del té verde. Mientres años estudióse'l so efeutu nel home, y apocayá (5 de marzu de 2007[22]) concluyóse que tienen un efeutu mínimu o nulu nel organismu humanu como antioxidantes.
Por eses causes son prescriptas les dietes riques en flavonoides, atópase en toles verdures pero les concentraciones más importantes pueden atopase nel brócoli,[21] la soya, el té verde y negru, el vinu, y tamién pueden inxerise en dellos suplementos nutricionales, xunto con ciertes vitamines y minerales. Nos frutos, les mayores concentraciones atópase na piel, polo que ye meyor comelos ensin pulgar, debidamente llavaos primeramente.[17] Tamién ye importante destacar que munchos d'estos compuestos atópase en proporciones variables nos distintos tipos de vinos, siendo responsables del efeutu preventivu que tien el consumu moderáu de vinu sobre les enfermedaes cardiovasculares, cáncer y otres enfermedaes dexeneratives.[8] Les mayores concentraciones nel tomate tán presentes nel de tipu "cherry", y na llechuga, na del tipu "Lollo Rosso".[23] La concentración de los flavonoides tamién varia enforma ente plantes de la mesma especie, polo que s'encamienta'l consumu de verdures de bona calidá, y como los flavonoides estropiar con facilidá, ye encamentáu consumiles no posible crudes, y si cocínense nun s'encamienta l'usu del microondes nin conxelales antes de ferveles,[23]
Aplicaciones nes plantes ornamentales y los frutos comerciales
Les plantes de mayor valor estéticu tienen mayor demanda nel mercáu, polo que s'aplicaron les conocencies en flavonoides pa realzar les coloraciones de les plantes.
La fenilalanina amonioliasa ye activada por situaciones de estrés, como pue ser el fríu. A baxes temperatures munches plantes, como les orquídees, pueden presentar coloraciones acolorataes (o violetes) en fueyes que primeramente yeren verdes. Esto ye nun ye más qu'un mecanismu de defensa, pero puede tener aplicaciones comerciales: si pulverizar con agua un frutu nes hores de mayor temperatura, favorezse la coloración nes zones en que nun se desenvuelve bien el color. Esti tratamientu produz, al evaporase l'agua, un descensu de temperatura del frutu d'hasta 10º C, lo que frenaría temporalmente les reaiciones de respiración y favorecería la reaición de síntesis de cianidina produciendo unos frutos de colores más vivos, de más valor comercial.
Investigación en flavonoides
La investigación en flavonoides puede estremase en dos grandes cañes: la investigación básica, que s'ocupa de resolver les víes biosintéticas de los flavonoides y los sos mecanismos de regulación, y l'investigación aplicada, representada pola inxeniería xenética de flavonoides, que manipolia l'ADN de les plantes con fines comerciales. Les dos cañes interrelacionar de forma estrecha, de forma que per un sitiu, ye necesariu conocer les víes biosintéticas y de regulación por que la manipulación del ADN tenga meyores probabilidaes d'ésitu, y per otru llau, la mesma manipulación del ADN refundia dacuando resultaos qu'ayuden a entender les víes biosintéticas y de regulación. Les dos cañes estrémense principalmente nos oxetivos propuestos, pero asolapar en dalgunos de los métodos utilizaos.
Un tercer aspeutu de la investigación ye l'usu de les víes biosintéticas y de regulación de los flavonoides como ferramienta pa esperimentación n'otres árees, y l'usu de los flavonoides en sí mesmos, como ferramienta pa estremar les especies en Botánica Sistemática.
Investigación de les víes biosintéticas y mecanismos de regulación
Les víes biosintéticas básiques fueron resueltes al traviés de la cuidadosa identificación y carauterización de numberoses enzimes qu'intervienen na biosíntesis. Nesta etapa de la investigación, aprovechóse la utilidá de munchos texíos de plantes que teníen la carauterística de tener enzimes de la síntesis de flavonoides en grandes cantidaes y que podíen ser aisllaes con facilidá. Exemplos de trabayos d'esti tipu son les célules irradiaes de perexil rizáu (Petroselinum hortense) de les que s'aislló la chalcona sintasa (Kreuzaler et al. 1979[24]); los cultivos en suspensión de célules de granes de soya (Glycine max) y poroto (Phaseolus vulgaris) de los que s'aislló la chalcona isomerasa (Moustafa y Wong 1967[25] Dixon et al. 1982,[26]); y les flores de Matthiola incana, petunia (Petunia hybrida), y Dianthus caryophyllus, de les que s'aislló la flavanona 3-hidroxilasa, la flavonol sintasa, la flavonoide 3'-hidroxilasa, y la dihidroflavonol reductasa (Forkmann et al. 1980[27] Spribille y Forkmann 1984,[28] Britsch y Grisebach 1986,[29] Stich et al. 1992,[30]). Estos esperimentos dexaron al descubiertu que les reaiciones químiques que mediaben na biosíntesis de flavonoides en plantes yeren una rede complexa, y propulsaron los esfuercios p'aisllar los correspondientes xenes que codificaban pa les enzimes descubiertes.[2]
Históricamente la clonación de xenes de la vía de los flavonoides amuesa qu'a mayoría de los xenes que codifican pa enzimes de la vía principal fueron primeru aisllaos per víes bioquímiques, por casu, por información estrayida direutamente de les carauterístiques de la enzima, o l'usu d'anticuerpos usando como antígeno a la enzima purificada. Tamién fueron útiles los mutantes que resultaben del insertamientu d'elementos transponibles nos xenes implicaos na vía biosintética (procedimientu llamáu "etiquetáu por transposones" utilizáu per primer vegada na historia de la ciencia por Federoff et al. en 1984[31] col qu'afayó'l xen que codificaba pa una enzima de la vía de los flavonoides). La clonación de xenes de la vía de la biosíntesis fíxose mayormente en maíz y petunia, con pocos apurras d'otres plantes. La vía de los isoflavonoides, que ta presente nes llegumes, foi estudiada apocayá en soya y alfalfa (Medicago sativa), magar los resultaos fueron principalmente basaos n'aproximamientos bioquímicos. Arabidopsis apaeció un pocu tarde na escena d'identificación de xenes, y fueron útiles principalmente los mutantes, yá que s'atoparon mutantes de Arabidopsis de la mayor parte de los xenes implicaos na vía biosintética de los flavonoides. L'usu de Arabidopsis ta enllenando vacíos de conocencia, por casu apocayá identificóse un xen que puede tar implicáu na síntesis de taninos entestaos (Devic et al. 1999.[32]). Al inxertar xenes de maíz nos mutantes de Arabidopsis comprobóse que les enzimes de la vía biosintética de los flavonoides caltuviéronse na so función en distancies evolutives enormes (Dong et al. 2001[33]). Los etiquetaos por transposón y por T-DNA en maíz, petunia y Arabidopsis tamién aproven información largamente esperada sobre los xenes envueltos nel tresporte de flavonoides del sitiu de síntesis na citoplasma hasta la vacuola (Marrs et al. 1995, Alfenito et al. 1998, Debeaujon et al. 2001).
La identificación de xenes que codifican pa factores reguladores ye más recién, y basóse cuasi puramente nel etiquetáu de transposones primeru, y l'etiquetáu de T-DNA dempués. Esto ye debíu principalmente a que les proteínes reguladores nun s'atropen en cantidaes importantes en nengún texíu, como sí lo faen les de biosíntesis, polo tanto nun son fáciles de realizar los aproximamientos de tipu bioquímicu con elles. Tamién hubo problemes al faer homoloxíes ente especies, porque s'atoparon secuencies altamente calteníes ente factores de trescripción (por casu los dominios bHLH y myb). Pero una vegada desenvolvióse l'etiquetáu por transposones, rápido aislláronse los factores reguladores de biosíntesis de flavonoides en maíz, petunia y boca de dragón (Antirrhinum majus). Factores reguladores adicionales fueron aisllaos darréu en Arabidopsis por clonáu posicional ("positional cloning") y etiquetáu de T-DNA. Un aproximamientu distintu foi realizada aisllando los factores de trescripción del perexil al traviés de South-western y screening de doble híbridu ("two-hybrid screening", Weisshaar et al. 1991, Rügner et al. 2001). L'usu de plantes trexéniques pa identificar y carauterizar los factores de regulación atoparon delles semeyances, pero tamién importantes diferencies, nos mecanismos polos que la vía de los flavonoides ye regulada en distintes especies de plantes (Lloyd et al. 1992, Quattrocchio et al. 1998, Uimari y Strommer 1998, Bradley et al. 1999).
La carauterización de la vía de los isoflavonoides n'alfalfa y soya hai provistu ferramientes pa la inxeniería metabólica de la síntesis de isoflavonoides n'otres especies de lleguminoses[34]
L'estudiu de les 3-desoxiantocianinas echó daqué de lluz sobre les sos víes biosintéticas (ver enllaz).
Tamién s'afayó un importante rol de los flavonoides na fertilidá masculina (fertilidá del polen) al traviés del analís de mutantes de maíz y de petunia que teníen una mutación na primer enzima de la biosíntesis de flavonoides. Sicasí un mutante de Arabidopsis na mesma enzima yera totalmente fértil, lo que demuestra que los flavonoides nun son riquíos por toles plantes pa la formación del tubu polínicu (pa review, ver Shirley 1996,[35]).
Tamién los flavonoides ayudaron a definir la especificidá de güéspede de microbios como Rhizobium spp. y Agrobacterium spp. (Rolfe 1988[36] Zerback et al. 1989,[37]).
Los flavonoides tamién contribúin al reconocencia de llantar güéspede per parte de les plantes parásites como Triphysaria versicolor y Cuscuta subinclusa, pero aparentemente nun son riquíos pal parasitismu esitosu de Arabidopsis per parte de Orobanche aegyptiaca (Kelly 1990[38] Albrecht et al. 1999,[39] Westwood 2000.[40]).
Pero según los flavonoides cumplen con funciones específiques n'especies distintes, tamién tienen una serie de roles que se caltienen llargamente, pa ello son útiles los modelos como Arabidopsis, qu'aprove información xenética y molecular que nun ta disponible n'otres plantes. Arabidopsis amás tien la ventaya de faer más simple la carauterización de la vía de los flavonoides, porque tien un solu xen pa la mayoría de les enzimes qu'intervienen na vía, a diferencia de lo que pasa con munches otres plantes que tienen munches copies d'esos xenes. Polo tanto una mutación nun xen de la vía ataya tol fluxu de la biosíntesis, en tolos texíos y en toles condiciones ambientales. Un exemplu de la so utilidá foi l'usu de dellos mutantes de Arabidopsis pa demostrar el rol inequívocu de los flavonoides en protexer a la planta de la radiación UV (Li et al. 1993[41]). Estos mutantes tamién aprovieron información sobre la contribución de los flavonoides presentes na cubierta de la grana pa caltener la dormición de la mesma (Debeaujon et al. 2000[42]). Ye más, la largamente revesosa hipótesis de que los flavonoides funcionaben como tresporte de la hormona auxina (Jacobs y Rubery 1988[43]) recibió sofitu d'estudios en Arabidopsis (Brown et al. 2001[44]). En cada casu Arabidopsis ayudó a dar les ferramientes pa investigar estos mecanismos n'otres especies de plantes.
Inxeniería xenética de flavonoides
- Ver tamién: En busca de la rosa azul
Por cuenta de les importantes funciones metabóliques que los flavonoides tienen nes plantes, les sos víes biosintéticas tán puramente regulaes. La inxeniería xenética aprovechó esta carauterística de los flavonoides pa faelos blancu de munchos trabayos d'inxeniería metabólica. La inxeniería metabólica de flavonoides puede definise como la teunoloxía que manipolia l'ADN qu'intervien na biosíntesis de flavonoides. L'ADN qu'intervien na biosíntesis de flavonoides puede estremase nel que codifica para compuestos estructurales y nel que codifica pa proteínes que regulen la trescripción (apocayá haise descripto que la biosíntesis puede ser regulada al nivel de la trescripción[45]).
La inxeniería metabólica de flavonoides empezó en 1987[46] y foi una área d'investigación bien granible na década del '90[47] Munchos de los procedimientos tán baxu patente.
La inxeniería xenética ye cara y costosa, polo que tienen de realizase primeramente esperimentos destinaos a conocer en fondura les víes metabóliques de la planta a tratar, coles mires de maximizar les probabilidaes d'ésitu del tratamientu. Por casu pueden deducise dellos puntos de les sos víes biosintéticas analizando la reaición de llantar ante la presencia de ciertos flavonoides conocíos.
Dellos exemplos d'aplicación esitosa de la inxeniería metabólica a los flavonoides son (Forkmann y Martens 2001.[48]):
- Coloración de les flores. La innovación nel color de les flores de les plantes ornamentales, cuantimás la ufierta de variedaes azules o marielles inesistentes na naturaleza, ye unu de los mayores curiosos que pueden ufiertar los cultivares. Como exemplu puede mentase la Petunia de flores naranxes (el color ye concedíu por flavonoides sintetizaos gracies a xenes traíos d'otres plantes), y los claveles violetes (llograos por xenes traíos de la petunia trexénica).
- Meyores nel potencial nutricional de los alimentos. Por casu nel tomate, llogróse introducir unos xenes del maíz qu'aumenten la biosíntesis de kaempferol en más d'un 60%, principalmente na magaya. Tamién s'introdució un xen de Petunia qu'aumenta la biosíntesis de la quercetina en más d'un 70 %, principalmente na piel. Tamién hai exemplos d'aplicación de la inxeniería en papes y en delles forrajeras.
- Meyores nel potencial farmacéuticu de les plantes. Dellos flavonoides presentes nes lleguminoses, llamaos isoflavonoides, pueden actuar como fitoestrógenos, lo cual xeneró interés nel usu d'estos compuestos pa tratar desordes hormonales n'humanos. La inxeniería xenética utilizar pa buscar la biosíntesis de isoflavonoides en plantes de cultivu onde de normal tán ausentes. Realizóse un esperimentu inicial esitosu en Arabidopsis thaliana.
- Supresión de la fertilidá del polen. En granes d'híbridos como'l maíz utilizóse la inxeniería xenética por que'l polen xeneráu por estes plantes híbrides fuera manera. Por casu nel maíz llogróse crackeando dos xenes CHS: C2 y Whp, lo que da como resultáu polen maneru de color blancu, productu de l'ausencia de flavonoides nél. Tamién hai exemplos n'híbridos de petunies y de la planta del tabacu.
- Biosíntesis de flavonoides por bacteries xenéticamente modificaes. Apocayá por aciu la inxeniería xenética llogróse cultivar bacteries capaces de sintetizar flavonoides de tipu flavanonas[49]
Usu de los flavonoides pa esperimentación n'otres árees
Los flavonoides o les sos víes biosintéticas fueron tamién utilizaos pa esperimentar n'otres árees de la ciencia, por casu:
Usu de les víes biosintéticas de los flavonoides como ferramienta. Los flavonoides contribuyeron en forma direuta o indireuta nel descubrimientu de munchos principios biolóxicos nos postreros 150 años.[2] Dos exemplos bien conocíos son l'usu que Mendel dio-y a los colores de les flores y les granes de Pisum sativum, ente otros calteres, pa desenvolver les sos teoríes alrodiu de los mecanismos del heriedu; y l'estudiu de la pigmentación de los granos de maíz de Barbara McClintock que llevó al descubrimientu de los elementos móviles nel ADN.
Más apocayá, los analises de la pigmentación nel maíz y los sos texíos vexetativos identificaron el fenómenu epixenéticu conocíu como paramutación, nel que les interaiciones ente alelos resulten en cambeos heredables na espresión xenética (Chandler et al. 2000[50]). De manera similar, los efeutos de la espresión de flavonoides trexénicos na pigmentación de la flor de petunia dexó al descubiertu'l fenómenu de cosupresión (Que y Jorgensen 1998[51] Metzlaff et al. 2000,[52]).
La vía de los flavonoides tamién foi un suxetu d'interés pa los estudios d'evoluciónsobremanera na Ipomoea purpurea, qu'ufierta recursos xenéticos únicos y una llarga historia d'analís (Iida et al. 1999[11] Rausher et al. 1999,[12] Durbin et al. 2000.[13]). Estos estudios sofiten la idea de que les enzimes de la biosíntesis de los flavonoides fueron derivaes d'enzimes del metabolismu primariu, y que la duplicación de xenes dexó l'adautación d'eses enzimes a funciones específiques.
Amás, la vía de los flavonoides, y la vía del fenilpropanoide de la que sale, tán sirviendo de modelos esperimentales pa entender la organización intracelular del metabolismu, con unos trabayos recién n'alfalfa y Arabidopsis qu'aproven información nueva na canalización d'intermediarios (channeling of intermediates) y nel asemblaje de complexos multienzimáticos (ver review en Winkel-Shirley 2001[1]).
Usu de los flavonoides en Botánica Sistemática. La Botánica Sistemática ye la ciencia que s'ocupa d'establecer rellaciones de parentescu ente les plantes a partir de les sos carauterístiques morfolóxiques, anatómiques, fisiolóxiques, la so estructura del ADN, etc. La Botánica Sistemática asume que mientres más asemeyaes son dos plantes ente sigo, más probable ye que tean cercanamente emparentaes. Por eso mientres más carauterístiques a analizar haya, más precisa va ser la determinación del parentescu.
Los flavonoides son estensamente utilizaos en Botánica Sistemática, probablemente porque son fáciles d'estrayer ya identificar[10] Por cuenta de que son bien variables, son más útiles en determinar rellaciones ente especies cercanamente emparentaes (o inclusive n'estudios de variación ente poblaciones de la mesma especie), pero tamién son dacuando útiles pa determinar rellaciones filoxenétiques a niveles más altos (Bate-Smith 1968,[53] Crawford 1978,[54] Gornall et al. 1979,[55] Harborne y Turner 1984,[56]). Finalmente, la diversidá na estructura química de los flavonoides demostró ser útil n'estudios d'hibridación ente especies (ver Alston y Turner 1963[57] Smith y Levin 1963,[58] Crawford y Giannasi 1982[59]).
El papel que xugaron los flavonoides n'establecer rellaciones filoxenétiques ye induldable. Por casu, la presencia de ciertos 5-desoxiflavonoides nes plantes del xéneru Amphypteryngium (que usualmente fuera allugáu na so propia familia, Julianaceae, por casu en Cronquist 1981) sofita'l so allugamientu nes anacardiáceas.
Síntesis
- Síntesis de Auwers
Referencies
- 1 2 3 Winkel-Shirley, B. 2001. "Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology". Plant Physiology 126: 485-493.
- 1 2 3 Winkel-Shirley, B. 2001b. "It takes a garden. How work on diverse plant species has contributed to an understanding of flavonoid metabolism". Plant Physiology 127: 1399-1404. (pdf equí)
- ↑ Flavonoides (isoflavonoides y neoflavonoides). IUPAC Compendium of Chemical Terminology. (pdf equí.
- ↑ Martínez-Flórez S., J. Gonález-Gallego, J. M. Culiebres y M. J. Tuñón. 2002. "Los flavonoides: propiedaes y aiciones antioxidantes". Nutr Hosp 17: 271-278. (pdf equí)
- ↑ Williams, CA, Grayer, RJ. 2004. "Anthocyanins and other flavonoids". Nat. Prod. Rep. 21: 539-573. (
- ↑ Major Types Of Chemical Compounds In Plants & Animals: Part II Flavonoids
- ↑ Trabayu práuticu sobre la flor Trabayo Práuticu sobre la flor
- 1 2 3 4 5 J. Palazón, R.M. Cusidó y C. Morales Metabolismu y significación biolóxica de los polifenoles del vinu, Grupu de Bioteunoloxía Vexetal, Facultá de Farmacia, Universidá de Barcelona.
- ↑ Mónica Noel Sánchez González y John P.N. Rosazza, Conversión de Chalconas a Auronas por Aspergillus alliaceus UI315 Resume
- 1 2 Judd, W. S. Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. 2002. "Secondary Plant Compounds" en: Plant systematics: a phylogenetic approach, Second Edition. Sinauer Axxoc, USA. Capítulu 4.
- 1 2 Iida S, A Hoshino, Y Johzuka-Hisatomi, Y Habu y Y Inagaki. 1999. "loricultural Traits and Transposable Elements in the Japanese and Common Morning Gloriesa". Annals of the New York Academy of Sciences 870: 265-274 (resume y pdf equí).
- 1 2 Rausher MD, RE Miller, P Tiffin. 1999. "Patterns of evolutionary rate variation among genes of the anthocyanin biosynthetic pathway". Molecular Biology and Evolution 16: 266-274 (resume y pdf equí).
- 1 2 Durbin ML, B McCaig, MT Clegg. 2000. "Molecular evolution of the chalcone synthase multigene family in the morning glory genome". Plant Molecular Biology 42: 79-92. (resume equí)
- ↑ Winkey-Shirley, B. 2001. "It takes a Garden. How work on diverse plant species has contributed to an understanding of flavonoid metabolism". Plant Physiology 127: 1399-1404.
- ↑ Kimura Y., T. Aoki, S. Ayae. 2001. "Chalcone isomerase isozimes with different substrate specifities towards 6'-hydroxy and 6'-deoxychalcones in cultured cells of Glycyrrhiza echinata, a leguminous plant producing 5-deoxyflavonoids". Plant Cell Physiol 42: 1169-1173.
- ↑ Brown D. Y., A. M. Rashotte, A. S. Murphy, J. Normanly, B. W. Tague, W. A. Peer, L. Taiz, G. K. Muday. 2001. "Flavonoids act as negative regulators of auxin transport in vivo in Arabidopsis thaliana". Plant Physiol 126: 524-535.
- 1 2 Principios activos de les plantes melecinales: los flavonoides Botanical Online
- ↑ Graf BA, Milbury PE, Blumberg, JB: "Flavonols, flavones, flavanones, and human health: epidemiological evidence." J Med Food 8: 281–290 (resume equí).
- ↑ Noticia: Curen el cáncer de fégadu nun esperimentu con consumu de quercetina (Noticia de 20 minutos)
- ↑ M. G. Hertog et al. 1995. "Flavonoid intake and long-term risk of coronary heart disease and cancer in the seven countries study". Archives of Internal Medicine Vol. 155 Non. 4. (resume equí, ver tamién la llista d'artículos que lu citen)
- 1 2 Yochum, L. et al. 1999. "Dietary Flavonoid Intake and Risk of Cardiovascular Disease in Postmenopausal Women." American Journal of Epidemiology 149:10 (resume equí)
- ↑ Balz Frei. 5 de marzu de 2007. "Studies force new view on biology of flavonoids". EurekAlert! (testu online equí).
- 1 2 Crozier, A. et al. 1997. "Quantitative Analysis of the Flavonoid Content of Commercial Tomatoes, Onions, Lettuce, and Celery" J. Agric. Food Chem., 45 (3), 590 -595. (resume equí)
- ↑ Kreuzaler F, Ragg H, Heller W, Tesch R, Witt I, Hammer D, Hahlbrock K. "Flavanone synthase from Petroselinum hortense. Molecular weight, subunit composition, size of messenger RNA, and absence of pantetheinyl residue." Eur J Biochem. 1979 99(1): 89–96. (resume equí)
- ↑ Moustafa, Y. y Wong, Y. 1967. "Purification and properties of chalcone-flavonone isomerase from soya bean seed." Phytochemistry 6: 625-632.
- ↑ Dixon RA, Dey PM, Whitehead IM. 1982. "Purification and properties of chalcone isomerase from cell suspension cultures of Phaseolus vulgaris". Biochim. Biophys. Acta 715: 25-33.
- ↑ Forkmann, G., Heller, W. y Grisebach, H. "Anthocyanin biosynthesis in flowers of Matthiola incana flavanone 3- and flavonoid 3'-hydroxylases." Z. Naturforsch. 35: 691-695
- ↑ Spribille, R. y Forkmann, G. 1984. "Conversion of dihydroflavonols to flavonols with enzyme extracts from flower buds of Matthiola incana". Z. Naturforsch. 39: 714-719.
- ↑ Britsch L y Grisebach H. 1986. "Purification and characterization of (2S)-flavanone 3-hydroxylase from Petunia hybrida". European Journal of Biochemistry 156: 569-577
- ↑ Stich K, Eidenberger T, Wurst F, Forkmann G. 1992. "Enzymatic conversion of dihydroflavonols to flavan-3,4-diols using flower extracts of Dianthus caryophyllus L.(Carnation)". Planta 187: 103–108.
- ↑ Federoff, N. V., Furtek, D. B. y Nelson, O. Y. "Cloning of the Bronze Locus in Maize by a Simple and Generalizable Procedure Using the Transposable Controlling Element Activator (Ac)" Proc. natn. Acad. Sci. U.S.A. 81: 3825−3829. (resume y pdf equí)
- ↑ Devic M, J Guilleminot, I Debeaujon, N Bechtold, Y Bensaude, M Koornneef, G Pelletier, M Delseny. 1999. "The BANYULS gene encodes a DFR-like protein and is a marker of early seed coat development" The Plant Journal 19 (4), 387–398. (resume y pdf equí)
- ↑ X Dong, EL Braun, Y Grotewold. 2001. "Functional Conservation of Plant Secondary Metabolic Enzymes Revealed by Complementation of Arabidopsis Flavonoid Mutants with Maize Xenes". Plant Physiology 127: 46-57 (resume y pdf equí).
- ↑ review en Dixon y Steele 1999
- ↑ Shirley, BW. 1996. "Flavonoid biosynthesis:'new' functions for an 'old' pathway" Trends Plant Sci 1:377-382.
- ↑ Rolfe BG. 1988. "Flavones and isoflavones as inducing substances of legume nodulation." Biofactors 1: 3-10 (resume equí)
- ↑ R Zerback, K Dressler, D Hess. 1989. "Flavonoid compounds from pollen and stigma of Petunia hybrida: inducers of the vir region of the Agrobacterium tumefaciens Ti plasmid". Plant Sci 62: 83-91.
- ↑ Kelly CK. 1990. " Plant Foraging: A Marxinal Value Model and Coiling Response in Cuscuta Subinclusa". Ecology 71: 1916-1925 (resume equí).
- ↑ Albrecht H, JI Yoder, DA Phillips. 1999. "Flavonoids Promote Haustoria Formation in the Root Parasite Triphysaria versicolor" Plant Physiology 119: 585-591. (resume y pdf equí)
- ↑ Westwood JH. "Characterization of the Orobanche–Arabidopsis system for studying parasite–host interactions" Weed Science 48: 742-748 (resume equí).
- ↑ Li J, TM Ou-Lee, R Raba, RG Amundson, RL Last. 1993. "Arabidopsis Flavonoid Mutants Are Hypersensitive to UV-B Irradiation". The Plant Cell 5: 171-179 (resume y pdf equí)
- ↑ Debeaujon I, M Koornneef. 2000. "Gibberellin Requirement for Arabidopsis Seed Germination Is Determined Both by Testa Characteristics and Embryonic Abscisic Acid". Plant Physiology 122: 415-424 (resume y pdf equí).
- ↑ Jacobs M, PH Rubery. "Naturally Occurring Auxin Transport Regulators". Science 241: 346-349 (resume equí).
- ↑ Brown DE, AM Rashotte, AS Murphy, J Normanly, BW Tague, WA Peer, L Taiz y GK Muday. 2001. Plant Physiology 126: 524-535 (resume y pdf equí)
- ↑ Nesi N., C. Jond, I. Debeaujon, M. Caboche, L. Lepiniec. 2001. "The Arabidopsis TT2 gene encodes an R2R3 MYB domain protein that acts as a key determinant for accumulation in developing seed". Plant Cell 13: 2099-2114.
- ↑ Meyer P, Heidmann I, Forkmann G, Saedler H. 1987. "A new Petunia flower colour generated by transformation of a mutant with a maize gene". Nature 330: 667-668.
- ↑ Dixon R, Steele C. 1999. "Flavonoids and isoflavonoids - a gold mine for metabolic engineering". Trends Plant Sci 4: 394-400.
- ↑ Forkmann, G., S. Martens. 2001. "Metabolic engineering and applications of flavonoids". Current Opinion in Biotechnology 12: 155–160 (pdf equí).
- ↑ Hwang Y. I. , M. Kaneko , Y. Ohnishi , S. Horinouchi. 2003. "Production of plant-specific flavanones by Escherichia coli containing an artificial gene cluster." Appl Environ Microbiol. 69 (5): 2699-706
- ↑ Chandler VL, WB Eggleston, JE Dorweiler. 2000. "Paramutation in maize". Plant Molecular Biology 43: 121–145. (resume equí)
- ↑ Que Q, RA Jorgensen. 1998. "Homology-based control of gene expression patterns in transgenic petunia flowers". Developmental Genetics 22: 100-109 (resume equí).
- ↑ Metzlaff M, M O'dell, R Hellens, RB Flavell. 2000. "Developmentally and transgene regulated nuclear processing of primary transcripts of chalcone synthase A in petunia" The Plant Journal 23: 63–72. (resume equí).
- ↑ Bate-Smith, Y. C. 1968. "The phenolic constituents of plants and their taxonomic significance" J. Linnean Soc. Bot. 60: 325-383.
- ↑ Crawford, D. J. 1978. "Flavonoid chemistry and angiosperm evolution." Bot. Rev. 44: 431-456.
- ↑ Gornall, R. J., B. A. Bohm y R. Dahlgre. 1979. "The distribution of flavonoids in the angiosperms". Bot. Notiser. 132: 1-30.
- ↑ Harborne, J. B. y B. L. Turner. 1984. Plant chemosystematics. Academic Press, Londres.
- ↑ Alston, R. Y. y B. L. Turner. 1963. "Natural hybridization among four species of Baptisia (Leguminosae)". Am. J. Bot. 50: 159-173.
- ↑ Smith, D. M. y D. A. Levin. 1963. "A chromatographic study of reticulate evolution in the Appalachian Asplanium complex". Am. J. Bot. 50: 952-958.
- ↑ Crawford, D. J. y D. Y. Giannasi. 1982. "Plant chemosystematics". BioScience. 32: 114-118, 123-124.
Bibliografía y enllaces
- Bibliografía
- Bioteunoloxía nel Cultivu d'Especies Ornamentales. Escandón, Alejandro S., Bioteunoloxía y Meyoramientu Vexetal páxs. 255-266
- Flavonoides, isoflavonoides y salú. María Rosario de Felipe y José Manuel Pozuelo, Centru de Ciencies Medioambientales (CSIC), Schironia Nº 3- Xunetu de 2004
- AN9, a Petunia Glutathione S-Transferase Required for Anthocyanin Sequestration, Is a Flavonoid-Binding Protein. Lukas A. Mueller, Christopher D. Goodman, Rebecca A. Silady, and Virginia Walbot, Plant Physiology, August 2000, Vol. 123, páxs. 1561–1570,
- Xeneral Plant Metabolism. Organised by N. Smirnoff for the Plant Metabolism Group, Abstracts / Comparative Biochemistry and Physiology Part A 132 (2002) S173–S180
- The characterisation of New Zealand Podocarpus hybrids using flavonoid markers. ROSEMARY F. WEBBY, KENNETH R. MARKHAM, New Zealand Journal of Botany, 1987, Vol. 25: 355-366
- The Unique Occurrence of the Flavone Aglycone Tricetin in Myrtaceae Pollen. Maria G. Camposa, Rosemary F. Webbyb and Kenneth R. Markhamb, 0939Ð5075/2002/0900Ð0944 $ 06.00 ” 2002 Verlag der Zeitschrift für Naturforschung, Tübingen
- Estudiu del efeutu solvatocrómico en derivaos fenólicos naturales. Romero Ale, Y; Olives, AI; Martín, L; Martín, MA;* Del Castillo, B.; Agnese, AM.; Ortega, MG; Núñez-Montoya, S; Cabrera JL*, Ars Pharmaceutica, 43:1-2; 57-71, 2002
- Cursu Popular de Cata de Vinos. 1997, Manuel Ruiz Hernández, Miguel Martínez Garoña
- Arabidopsis ICX1 Is a Negative Regulator of Several Pathways Regulating Flavonoid Biosynthesis Xenes. Helena K. Wade, Awinder K. Sohal2, and Gareth I. Jenkins, Plant Physiology, February 2003, Vol. 131, páxs. 707–715,
- Section Y SNIF part 2: Summary information format for products containing genetically modified higher plants (GMHPs). Florigene Moonlite (123.2.28)
- Enllaces bibliograficos
- Flavonoides
- Antocianinas - Papel indicador de Ph y estudo da estabilidade da solução de repolho roxo
- A Brief Introduction to the Floriculture Research Center, Agricultural Research Institute, Council of Agriculture
- Dihydroquercetin dimers by oxidative coupling reactions
- Annie's special plant lists
- Ecology & Evolutionary Biology Conservatory Bletilla striata
- Die lachsrote Petunia
- Policromía de les Palmes por Sven Nehlin - The Multicolored Palms
- Park Garden
Enllaces rellacionaos non bibliográficos
- Anthocyanins and other flavonoids. J. B. Harborne and C. A. Williams, Plant Science Laboratories, University of Reading, Reading, UK, RG6 6AS (Bien bonu pa buscar qué flavonoides presentar en determinaes plantes).
- Horse Chestnut (Aesculus hippocastanum). Mary McLellan, RN, BSN, Longwood Herbal Task Force, Revised June 15, 2000
- Nuevos colores en flores por aciu inxeniería xenética
- La Composición química de la xamasca
| Control d'autoridaes |
|
|---|
 Datos: Q3561192
Datos: Q3561192 Multimedia: Flavonoids
Multimedia: Flavonoids