Dimetrodon | ||
|---|---|---|
| Rangu temporal: 299 Ma-270 Ma Permianu Inferior | ||
 | ||
| Clasificación científica | ||
| Reinu: | Animalia | |
| Filu: | Chordata | |
| Clas: | Synapsida | |
| Orde: | Pelycosauria | |
| Suborde: | Eupelycosauria | |
| (ensin clasif.): | Sphenacodontia | |
| Familia: | Sphenacodontidae | |
| Xéneru: |
Dimetrodon Cope, 1878 | |
| Especie tipu | ||
|
Dimetrodon limbatus (Cope, 1877 [orixinalmente Clepsydrops limbatus]) | ||
| Especies | ||
| Sinonimia | ||
Bathyglyptus Case, 1911 | ||
| Consultes | ||
| [editar datos en Wikidata] | ||
Dimetrodon («diente de dos midíes») ye un xéneru estinguíu de sinápsidos pelicosaurios perteneciente a la familia Sphenacodontidae que les sos especies vivieron n'América del Norte y Europa mientres el Permianu Inferior ente hai alredor de 299 y 270 millones d'años tras.[1][2][3]
La carauterística más sobresaliente de Dimetrodon ye la enorme vela dorsal, similar a una aleta, que la so estructura basar n'escayos vertebrales allargaes. La función que pudo desempeñar posiblemente foi la d'estabilizar la columna vertebral p'ameyorar la locomoción o la de calecer y esfrecer el cuerpu como un sistema de termorregulación. Sicasí, estudios recién sostienen que la vela pudo ser inefectiva pa regular el calor corporal y pudo cumplir nel so llugar una función reproductiva mientres esplegar destinaos al apareyamientu.
Con seguridá yera unu de los superdepredadores del so tiempu y alimentábase de pexes y otros tetrápodos, incluyendo reptiles y anfibios. Tenía una postura cuadrúpeda y tenía un craniu grande y de perfil curvu con grandes dientes de distintu tamañu agospiaos en dambos maxilares. La mayoría de los fósiles de Dimetrodon fueron topaos nel suroeste d'Estaos Xuníos, cuantimás d'unos depósitos llamaos Llechos Coloraos de Texas y Oklahoma, y creíase que la distribución del xéneru acutar a América del Norte. Sicasí, apocayá describióse una especie nueva topada n'Europa (Alemaña), lo cual nun ye d'estrañar yá que a principios del Permianu América del Norte y Europa faíen parte del supercontinente Euramerica. El nome del xéneru utilizar por primer vegada en 1978 y dende entós describiéronse una ventena d'especies, de les cualos una docena considérense válides.
Dimetrodon na cultura popular confundir frecuentemente con un dinosauriu o dalgunu de los sos contemporáneos, pero en realidá vivió siquier 40 millones d'años antes de l'apaición del primer dinosauriu mientres el Triásicu. A pesar de tener apariencia de reptil atópase más rellacionáu colos mamíferos que con cualesquier de los reptiles modernos; sicasí, la familia a la que pertenez nun fai parte del llinaxe que dio orixe a los mamíferos vivientes.
Historia

Descripción d'Cope

Los primeros estudios realizaos en fósiles anguaño clasificaos como Dimetrodon realizar el paleontólogu estauxunidense Edward Drinker Cope na década de 1870. Cope llogró los fósiles xunto con munchos otros restos de tetrápodos del Permianu, de manes de dellos cazadores de fósiles que los llograron nos Llechos Coloraos de Texas. Ente los cazadores atopaba'l naturalista suizu Jacob Boll, el xeólogu estauxunidense W. F. Cummins y el paleontólogu aficionáu Charles Hazelius Sternberg.[4]
Sternberg unvió-y dalgunos de los sos fósiles al paleontólogu alemán Ferdinand Broili de la Universidá de Múnich, sicasí Broili nun foi tan prolíficu como Cope a la de describir los especímenes. El rival d'Cope Othniel Charles Marsh tamién llogró dellos fósiles de Dimetrodon qu'unvió al Muséu Walker.[5] El nome Dimetrodon utilizar por primer vegada en 1878 cuando Cope describió les especies Dimetrodon incisivus, Dimetrodon rectiformis y Dimetrodon gigas na publicación científica Proceedings of the American Philosophical Society.[6]
Sicasí, la primer descripción d'un fósil de Dimetrodon tuvo llugar un añu antes, cuando Cope describió la especie Clepsydrops limbatus proveniente de los Llechos Coloraos de Texas.[7] (El nome Clepsydrops foi acuñáu por Cope en 1875 pa describir esfenacodóntidos del Condáu de Vermilion (Illinois) y más tarde utilizáu pa nomar munchos especímenes de esfenacodóntidos de Texas; a finales del sieglu XIX y principios del sieglu XX munchos esfenacodóntidos de Texas asignáronse tanto a Clepsydrops como a Dimetrodon) C. limbatus foi revaluado como una especie de Dimetrodon en 1940, polo cual la publicación d'Cope de 1877 convertir nel primer rexistru de Dimetrodon.
El mesmu Cope foi l'encargáu de describir en 1978 la primer especie de sinápsido con vela dorsal, col nome de Clepsydrops natalis; sicasí consideró la vela como una aleta y comparar cola cresta de los actuales basiliscos (Basilicus). Les veles nun taben calteníes nos especímenes de D. incisivus y D. gigas descritos tamién na so publicación de 1878, pero si lo taben nel espécime de D. rectiformis tamién descritu por él.[6]
Descripciones d'entamos del sieglu XX

Nes primeres décades del sieglu XX, el paleontólogu estauxunidense Y. C. Case foi l'autor de munchos estudios sobre Dimetrodon y describió delles especies. Case recibió fondos del Institutu Carnegie pa estudiu de munchos especímenes de in coleiciones del Muséu Americanu d'Historia Natural y otros museos.[5] Munchos d'estos especímenes fueron topaos por Cope, pero nun los detalló bien a xeitu, yá que Cope yera conocíu por describir especies nueves basándose solo en dellos fragmentos óseos. A principios de la década de 1920, el paleontólogu Alfred Romer revisó munchos especímenes de Dimetrodon y describió otres especies. En 1940, Romer foi'l coautor d'un estudiu estensu xunto a Llewellyn Ivor Price que llamaron Review of the Pelycosauria, nel cual les especies de Dimetrodon descrites por Cope y Case fueron revaluadas.[8] La mayoría de les especies consideraes válides por Romer y Price guarden inda esi estáu.[9]
Nuevos especímenes
Nes décades posteriores a la monografía de Romer y Price afayáronse nuevos especímenes de Dimetrodon en llocalidaes fora de Texas y Oklahoma n'Estaos Xuníos. El primeru d'ellos foi afayáu na rexón de Cuatro Esquines Utah en 1966;[10] otru en Arizona en 1969,[11] y en 1975 n'Ohio.[12] Una especie nueva del xéneru denomada D. occidentalis foi descrita en 1977 procedente de Nuevu Méxicu.[13] Los fósiles topaos n'Utah y Arizona probablemente tamién pertenecíen a D. occidentalis.[14]
Antes d'estos afayos, formulóse una teoría que proponía la esistencia d'un pasu marítimu a principios del Permianu que dixebraba lo qu'anguaño son los estaos de Texas y Oklahoma de los territorios del oeste, acutando a les especies de Dimetrodon a una pequeña rexón d'América del Norte, ente que un esfenacodóntidos más pequeñu llamáu Sphenacodon apoderaba l'área occidental. A pesar qu'esta franxa d'océanu probablemente si esistía, el descubrimientu de fósiles fora d'esti territoriu amuesa que la so estensión yera llindada y nun constituyía una barrera efectiva pa la distribución de les especies de Dimetrodon.[13][15]
En 2002, una especie nueva de Dimetrodon que pasó a llamase D. teutonis describir a partir d'un afayu na Cuenca Saar-Nahe n'Alemaña, lo que constituyó'l primer afayu d'una especie d'esti xéneru fora d'América del Norte.[16]
Filoxenia
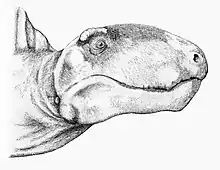
Dimetrodon ye un miembru primitivu dientro de los sinápsidos, un grupu diversu qu'inclúi los mamíferos vivientes y tolos sos parientes estinguíos. El xéneru pudo evolucionar d'una forma primitiva de la familia Sphenacodontidae, probablemente de Sphenacodon ferox mientres el Carboníferu Cimeru o'l Permianu Inferior (Gzheliense-Asseliense);[17] y la so desapaición coincide cola puxanza de los terápsidos eotitanosúquidos y dinocéfalos mientres el Ufimiano.[17] Na cultura popular con frecuencia confundir con un dinosauriu, a pesar que surdieron a lo menos 40 millones d'años primero que'l primer dinosauriu apaeciera.[18] Dimetrodon atópase más emparentáu colos mamíferos actuales que colos dinosaurios y cualesquier de los reptiles modernos. Dende finales del sieglu IXX hasta los entamos del sieglu XX, la mayoría de los paleontólogos considerar un reptil acordies cola taxonomía de Linneo. Nesta clasificación los reptiles fueron consideraos como una clase y Dimetrodon incluyóse como un xéneru d'esta clase. Sicasí, los paleontólogos reconocieron que los mamíferos evolucionaron d'esti grupu no qu'ellos llamaron una transición de reptil a mamíferu. Más apocayá, la taxonomía filoxenética unificó la clasificación de los vertebraos y definió que toos comparten un ancestru común. Los reptiles y mamíferos atópense allugaos en clados distintos, y cada grupu tien un ancestru común del cual toos son los sos descendientes. So la sistemática filoxenética, los descendientes del ancestru común más recién de Dimetrodon y tolos reptiles vivientes inclúi tamién a tolos mamíferos modernos y los sos antepasaos, los qu'anguaño se consideren más rellacionaos con Dimetrodon que con cualquier reptil viviente. Los descendientes del ancestru común más recién de los mamíferos y reptiles (el cual apaeció hai alredor de 310 millones d'años nel Carboníferu Cimeru) estremar en dos clados: los sinápsidos, que inclúi a Dimetrodon y a los mamíferos y los saurópsidos, qu'inclúin a los reptiles vivientes y tolos reptiles estinguíos más rellacionaos unos con otros que colos mamíferos.[2]

Dientro de los sinápsidos Dimetrodon fai parte del clado Sphenacodontia. Los esfenacodontos fueron clasificaos per primer vegada como sinápsidos primitivos en 1940 polos paleontólogos Alfred Romer y Llewellyn Ivor Price, xunto a los grupos Ophiacodontia y Edaphosauria.[8] Miembros d'estos trés grupos conocer del Carboníferu Cimeru y el Permianu Inferior. Romer y Price identificar principalmente por carauterístiques postcraneales como la forma de los miembros y les vértebres. Considerarona a Ophiacodontia el grupu más primitivu por causa de la so mayor semeyanza colos reptiles; mientres Sphenacodontia consideróse'l más avanzáu debíu'l so la mayor semeyanza colos terápsidos, que contienen a los sinápsidos más emparentaos colos mamíferos. Romer y Price asitiaron a una familia de sinápsidos primitivos llamada Varanopidae dientro de Sphenacodontia, considerándola más primitiva que los otros esfenacodóntidos rellacionaos con Dimetrodon.[19] Pensaron que los varanópidos y los esfenacodóntidos emparentaos con Dimetrodon taben estrechamente rellacionaos yá que dambos grupos yeren carnívoros; sicasí, los varanópidos yeren munchos más pequeños, escarecíen de vela dorsal y teníen aspeutu de reptil. La clasificación actual de les rellaciones ente sinápsidos foi propuesta pol paleontólogu Robert R. Reisz en 1986, que'l so estudiu basar principalmente nes carauterístiques craniales dexando en segundu planu'l restu de la cadarma.[20] Dimetrodon considérase un esfenacodontos so esta clasificación, pero los varanodóntidos son consideraos sinápsidos basales y clasifíquense fora de los esfenacodontos. Dientro de Sphenacodontia atópase'l clado Sphenacodontoidea, que de la mesma contién a la familia Sphenacodontidae y l'orde Therapsida. La familia Sphenacodontidae integra a Dimetrodon y otros sinápsidos dotaos de veles dorsales como Sphenacodon y Secodontosaurus, mientres los terápsidos inclúin a los mamíferos y la mayor parte de los sos parientes que vivieron mientres el Permianu y el Triásicu. De siguío represéntase un cladograma modificáu a partir del analís de Benson (en prensa) que sigue esta filoxenia:[19]
| Amniota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Descripción anatómica


Dimetrodon yera un sinápsido cuadrúpedu, posesor d'una gran vela con aspeutu d'aleta que-y cubría l'envés. El so tamañu y morfoloxía yera similar a un aligator modernu, quitando'l fechu de que se trataba d'un animal puramente terrestre. Dimetrodon foi'l depredador dominante del so ambiente por más de 25 millones d'años, tiempu mientres el cual diversificóse n'alredor d'una docena d'especies, qu'aumentaron el so tamañu col pasu del tiempu. La mayoría de les especies teníen un llargor que bazcuyaba ente 1,7 y 3,2 metros, y un pesu envaloráu que variaba ente 28 y 250 kg.[16] La especie más grande descrita de Dimetrodon ye D. grandis de 3,2 metros de llargor y la más pequeña ye D. teutonis de solu 60 cm.[21] Les especies grandes de Dimetrodon topar ente los mayores depredadores d'entamos del Permianu; sicasí, el so pariente Tappenosaurus, conocíu por afayos fósiles n'estratos daqué más recién, pudo ser un pocu más grande con un llargor envaloráu de 5,5 m.[22][23] Dientro de les carauterístiques que la identifiquen más colos mamíferos que colos reptiles cabo destacar la presencia d'un quexal inferior fundida, un dentame complexo pa mazcar, una anatomía muscular mandibular novedosa, un celebru más grande y trés pequeños güesos nel oyíu mediu.[18]
Craniu

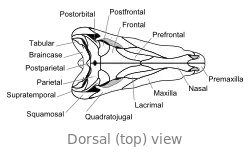
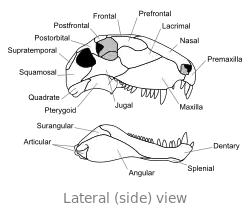
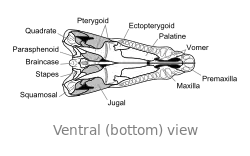
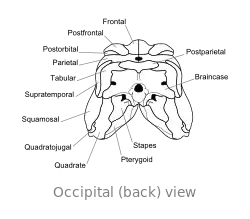
El craniu de Dimetrodon ye proporcionalmente alto, angostu y de perfil triangular vistu dende enriba. Los furos oculares asítiase na parte cimera y posterior del craniu. Posterior a cada cuenca ocular atópase un furacu llamáu fenestra infratemporal. Otru dos pares de furacos pueden reparase na cara posterior del craniu al trate dende enriba: la fenestra supratemporal y la fenestra postemporal. El polu posterior del craniu (occipuciu) empobinar nun ángulu llixeramente cimeru, una carauterística que comparte con otros sinápsidos primitivos.[24] Vistu lateralmente, la contorna cimera del craniu torna escontra baxo y alantre nun arcu convexu hasta la punta del focico. La punta del quexal cimeru, formada por güesu premaxilar, enllargar dende enriba sobre'l maxilar pa formar una discontinuidá, que forma un diastema ente los arcos dentales.
Pola so posición filoxenética tenía un patrón de musculatura mandibular a partir del cual anicióse'l de los mamíferos.[25] Sicasí, l'estudiu de la rexón temporal, el cielu la boca posterior y el quexal, indica que Dimetrodon tenía músculos aductores nel quexal con diferenciación y distribución similar a los reptiles modernos. Les carauterístiques de los músculos masticatorios y la presencia d'un aparatu prensil potente, suxuren que Dimetrodon alimentar d'animales grandes.[25]
Dientes
El llargor de los dientes difier notablemente d'alcuerdu al so allugamientu nos maxilares, a lo cual debe'l so nome Dimetrodon que significa «diente de dos midida», n'alusión a los dos xuegos de dientes de distintu llargor que tenía[26] Unu o dos pares de dientes similares a caninos esprender del maxilar superior y incisivos de gran tamañu proyeutar dende la punta de dambos maxilares insertos nel premaxilar y el quexal. Dientes más pequeños atópense alredor de la fienda maxilar y en direición posteriores a los caniniformes, faciéndose menores de manera progresiva escontra tras nos maxilares.[27]
Munchos de los dientes son más anchos na seición media y fáense más angostos al contautar col güesu maxilar, lo que-yos confier una forma ovalada o de «gota d'agua». Esta forma de los dientes ye carauterística de Dimetrodon y los esfenacodóntidos más rellacionaos, traza que dexa estremalos d'otros sinápsidos primitivos. Como n'otros sinápsidos basales, los dientes de la mayoría de les especies de Dimetrodon tienen cantos serruchaos. Los dientes de la especie más pequeña D. teutonis escarecen d'esti tipu de cantu, pero caltienen los sos cantos afilaos.[21]
Cuévanu nasal
Nos fósiles topaos, la superficie interna de la rexón nasal tien crestes llamaes nasoturbinales, que pudieron dar base al cartílagu que soportaba'l epiteliu olfatorio, que como'l so nome indica, encargar de detectar los golores. Estos riegos son muncho más pequeños que los presentes nos sinápsidos más recién del Permianu y Triásicu, nos cualos los grandes nasoturbinales considérense la prueba de que fueron animales de sangre caliente, por cuenta de que posiblemente sirvieron de soporte a la membrana mucosa que calez y amuga l'aire inspiráu. Por ello, el cuévanu nasal de Dimetrodon ye transicional ente los vertebraos terrestres primitivos y los mamíferos.[28]
Articulación mandibular y oyíu
Otra carauterística transicional de Dimetrodon ye una cresta nel envés del quexal llamáu llámina reflexa. Esta atópase nel güesu articular que coneuta'l maxilar inferior col güesu cuadráu del craniu pa formar l'articulación mandibular. Nos ancestros más recién de los mamíferos, el güesu articular y el cuadráu dixebrar de l'articulación mandibular, convirtiéndose l'articular nel martiellu del oyíu mediu. La llámina reflexa pasu a faer parte d'una estructura llamada aniellu timpánico que brinda soporte al tímpanu en tolos mamíferos vivientes.[29]
Cola

La cola de Dimetrodon constitúi una gran proporción de la so masa corporal y ta formada per alredor de 50 vértebres caudales. Nes primeres cadarmes descrites la cola taba ausente o incompleta; la única vértebra caudal conocida nun exemplar yera un númberu once, que s'atopaba al altor del cadril. Por cuenta de que los primeros afayos dexaben entever que les vértebres caudales menguaben rápido de tamañu al moviera escontra la punta de la cola, munchos paleontólogos del sieglu XIX y principios del sieglu XX pensaron que l'animal tenía una cola bien curtia. Esti conceutu camudó hasta 1927 cuando se describió un espécime con una cola cuasi completa.[30]
Vela

La carauterística más distintiva de Dimetrodon ye la gran vela con aspeutu d'aleta qu'ocupaba toa'l llargor del so envés. Esta formación anatómica aniciar a partir de la elongación de les apófisis espinosas de les vértebres. Cada escayu yera distintu na so seición tresversal, dende la base hasta la punta.[31] Na cercanía del cuerpu vertebral la seición tresversal del escayu tien una forma rectangular y escontra la punta toma una figura d'ocho con un riegu a cada unu de los sos llaos. Créese qu'esta forma dábalu más resistencia a los escayos lo que prevenía dobladures y quebres.[32] Una seición tresversal d'un escayu de Dimetrodon giganhomogenes tien forma rectangular pero caltién la configuración n'ocho escontra'l centru, indicando que la forma de los escayos puede camudar cola edá de los individuos.[33]
L'anatomía microscópica de cada escayu tamién varia dende la base hasta la punta ya indica los sitios que sirvíen d'asientu a los músculos del llombu y la porción que faía parte de l'aleta. La parte proximal del escayu tenía una superficie rugosa que pudo sirvir como puntu de anclaje pa los músculos dorsales. Tamién tenía una rede de texíu conectivo llamáu fibres de Sharpey qu'indiquen que s'atopaba enfiñíes dientro del cuerpu. Na parte distal de los escayos la superficie del güesu ye más llisa. El periostiu atópase cubiertu de pequeños riegos que probablemente sirvíen de canal a los vasos sanguíneos que irrigaban la vela.[9] Antes creíase que los grandes riegos llaterales de los escayos fueron canales derrotes por grandes vasos sanguíneos, pero por cuenta de que'l güesu escarez de canales vasculares de tamañu significativu camudóse'l conceutu y piénsase que la vela taba pocu vascularizada. En dellos especímenes de Dimetrodon pueden reparase árees deformadas que pueden ser quebres consolidaes. El güesu cortical sobre estes rotures atópase abondo vascularizado, lo que suxure que la vela cuntaba con texíos blandos que proyeutaben vasos sanguíneos sobre'l sitiu.[32] Capes concéntriques de güesu lamelar constitúin la mayor parte de los escayos al reparales nuna corte tresversal y contién zones de recesión de la crecedera que dexen calcular la edá de los especímenes al momentu de la so muerte.[34] En munchos afayos, la porción distal de los escayos xira sópito, indicando qu'en vida la vela pudo tener un perfil irregular. Esta combadura suxure que'l texíu blando pudo nun llegar hasta la punta de les proyeiciones ósees, lo cual significa que'l texíu ente los escayos nun yera tan estensu como se pensaba.[35]
Función termorreguladora

Esisten munches proposiciones alrodiu de la función de la vela dorsal nesti animal. Ente les primeres suxurióse que pudo sirvilu al animal pa camuflase ente los xuncos mientres acesmaba a la so presa o que pudo tratase d'una auténtica vela qu'impulsaba al animal sobre l'agua.[36] Otra posible función que pudieron tener los sos llargos escayos neurales foi la d'estabilizar el tueru por aciu la restricción de los sos movimientos verticales, lo que-y aprovía un meyor soporte pa los movimientos llaterales riquíos pa la marcha.[32] En 1940, Alfred Romer y Llewellyn Ivor Price propunxeron que la vela tenía una función termorreguladora, lo que-y dexaba al animal calecer el so cuerpu cola lluz solar. Nos años siguientes creáronse munchos modelos pa envalorar la capacidá de termorregulación de Dimetrodon. Por casu, nun artículu de 1973 de la revista Nature, los paleontólogos C. D. Bramwell y P. B. Fellgett envaloraron que lu tomaba a un animal de 200 kg cerca d'una hora y media p'amontar la so temperatura de 26 a 32 °C .[37] En 1986, Steven C. Haack concluyó que'l calentamientu yera más lentu de lo envalorao y que'l procesu tomaba alredor de cuatro hores. Por aciu l'usu d'un modelu basáu nuna variedá de factores ambientales y factores fisiolóxicos hipotéticos del animal, Haack atopó que l'aleta dexába-y a Dimetrodon calecer rápido na mañana y algamar una temperatura óptima mientres el día, pero yera inefectiva pa esaniciar l'escesu de calor y caltenese caliente mientres la nueche.[38] En 1999, un grupu d'inxenieros mecánicos escurrió un modelu virtual p'analizar la capacidá de la vela pa regular la temperatura corporal mientres los cambeos estacionales. Concluyeron que la vela yera útil pa prindar y lliberar el calor mientres tol añu.[39]
La mayoría d'estos estudios atribúin dos funciones termorreguladoras a la vela: una como un mediu de calecer rápido na mañana o otra como un mediu de menguar la temperatura corporal. Presumir de Dimetrodon, como de tolos vertebraos terrestres del entamu del Permianu, que se trataba d'un animal de sangre frío o ectotérmico, dependiente del sol pa caltener la temperatura corporal. Por cuenta del so gran tamañu, los cambeos de temperatura yeren más lentos que n'animales más pequeños. Al aumentar la temperatura na mañana, les preses de Dimetrodon, de menor tamañu, podía calecer y faese activos más rápidu que'l so depredador. Munchos paleontólogos, ente ellos Haack, propunxeron que la vela de Dimetrodon pudo dexa-y entrar en calor más rápido a l'alba, al son de les sos preses.[38] La gran superficie de l'aleta tamién pudo dexa-y esaniciar l'escesu de calor, por cuenta del metabolismu o de la radiación solar. Pa esfrecese, l'animal empobinar la so aleta paralela a la lluz solar o acutar el fluxu sanguíneo escontra l'aleta pa caltener el calor mientres la nueche.[36]

En 1986, J. Scott Turner y C. Richard Tracy propunxeron que'l desenvolvimientu de la vela en Dimetrodon ta rellacionáu cola evolución del sangre caliente nos ancestros de los mamíferos. Ellos pensaron que la esistencia de vela condució a esti animal escontra la homeotermia, al caltener constante, anque baxa, la temperatura corporal. Los mamíferos tamién son homeotérmicos, pero difieren de Dimetrodon por ser endotérmicos, yá que controlen la so temperatura corporal, ensin depender del mediu ambiente, por aciu l'aumentu del so metabolismu. Los mesmos autores notaron que los primeres terápsidos, un grupu avanzáu de sinápsidos estrechamente rellacionaos colos mamíferos, teníen miembros llargos que podríen estenar el calor de manera tan eficiente como l'aleta de Dimetrodon. La homeotermia desenvuelta nesti xéneru y los sos rellacionaos pudo conducir a los terápsidos a los cambeos de la so forma corporal, lo cual eventualmente desaguo nos mamíferos de sangre caliente.[40]
Los estudios más recién de la vela de Dimetrodon y otros esfenacodóntidos sofiten l'afirmación de Haack de 1986, de que la vela tenía una probe adautación pa lliberar calor o regular la temperatura corporal. La presencia de vela n'especies pequeñes como D. milleri y D. teutonis nun sofiten la idea de que la función de l'aleta fuera termorreguladora por cuenta de que les aletes pequeñes son menos efectives pa tresferir calor y un cuerpu pequeñu puede absorber y lliberar calor con facilidá ensin aditamentos adicionales. Amás, parientes cercanos de Dimetrodon como Sphenacodon teníen crestes dorsales pequeñes que con seguridá nun desempeñaben nenguna función termorreguladora.[9] Créese que les veles grandes de Dimetrodon evolucionaron a partir d'eses pequeñes crestes, lo cual significa qu'a lo llargo de la esistencia d'esta traza evolutiva, la termorregulación nun xugó un papel preponderante.[41]
Seleición sexual
Los especímenes más grandes de Dimetrodon tuvieron les veles más grande en proporción al so tamañu, un exemplu d'alometría positiva. Esta carauterística pudo ser benéfica pa la termorregulación, una y bones ello implica qu'ente más grande l'animal la superficie de la vela amóntase más rápidu que la so masa corporal. Los animales con cuerpos más grandes xeneren mayor cantidá de calor per mediu del metabolismu y el monto del calor que tienen d'estenar per mediu de la superficie del cuerpu ye sustancialmente mayor que nos animales de menor tamañu. La disipación efectiva de calor puede predicise en distintos animales d'una manera simple por aciu la rellación ente la masa y l'área de superficie corporal. Sicasí, un estudiu de 2010 de alometría en Dimetrodon atopó una rellación distinta ente la superficie de la vela y la so masa corporal: l'esponente real de la escala de la vela foi enforma mayor que l'esperáu nun animal afechu pa estenar el calor. Los investigadores concluyeron que la vela de Dimetrodon amosaba una tasa de crecedera más alta que la riquida pa cumplir una función termorreguladora y que la principal razón pa la evolución d'esta traza foi la seleición sexual.[41]
Dimorfismu sexual


Dimetrodon pudo tener dimorfismu sexual, lo que significa qu'esistía una discrepancia de tamañu ente los sexos. Dellos especímenes catalóguense como machos, ente otros, en base a la mayor espesura de los sos güesos, les veles y cranios más grandes y les fiendes maxilares más amplies. D'alcuerdu a estes diferencies les cadarmes del Muséu Americanu d'Historia Natural (AMNH 4636) y del Muséu Field d'Historia Natural pueden ser machos y la cadarma nel Muséu de Denver de Naturaleza y Ciencia (MCZ 1347) y el Muséu d'Historia Natural de la Universidá de Michigan pueden ser femes.[8]
Paleoecoloxía

Los fósiles de Dimetrodon conocer a partir d'afayos n'América del Norte (Estaos Xuníos) y Europa (Alemaña), rexones que faíen parte del supercontinente Euramérica a principios del Permianu. La mayor parte del material atribuyíu al xéneru vien de tres grupos xeolóxicos de los estaos de Texas y Oklahoma: el Grupu Clear Fork, el Grupu Wichita y el Grupu Pearce River.[42] La mayoría de los especímenes topaos fixeron parte d'ecosistemes de tierres baxes y planes mientres el Permianu, que pudieron ser grandes banzaos. N'especial, los Llechos Coloraos de Texas son una área con gran diversidá de fósiles de tetrápodos. Amás de Dimetrodon, los tetrápodos más comunes nos Llechos Coloraos y los xacimientos fósiles del Permianu Inferior nel sureste d'Estaos Xuníos, atópense los anfibios Archeria (animal), Diplocaulus, Eryops y Trimerorhachis, el reptilliomorfo Seymouria, el reptil Captorhinus y los sinápsidos Ophiacodon y Edaphosaurus. Estos tetrápodos faen parte d'un grupu d'animales que'l paleontólogu Everett C. Olson llamó «cronofauna del Permo-Carboníferu», que apoderó los ecosistemes del continente euramericano por dellos millones d'años.[43] Sobre la base de la xeoloxía de depósitos similares a los Llechos Coloraos, la redolada d'esta pudo tar apoderáu per tierres baxes con vexetación trupa tipu delta fluvial.[44]
Rede trófica
Olson fixo munches inferencias alrodiu de la paleoecoloxía de los Llechos Coloraos de Texas y el papel de Dimetrodon dientro d'esti ecosistema. Propunxo dellos tipos principales d'ecosistemes onde vivieron los primeres tetrápodos. Dimetrodon asignar al ecosistema más primitivu, que dependía de la rede alimenticia acuática. Nesti modelu, les plantes acuática funcionaben como'l productor primariu constituyendo la fonte d'alimentu pa los pexes ya invertebraos acuáticos. La mayor parte de los vertebraos terrestres alimentar d'estos comensales acuáticos. Dimetrodon probablemente yera un superdepredador nel so ecosistema, depredando una cantidá diversa d'organismos como'l tiburón primitivu Xenacanthus, los anfibios Trimerorhachis y Diplocaulus, y los tetrápodos terrestres Seymouria y Trematops. Conócense inseutos qu'habitaron esta zona que probablemente en dalgún grau faíen parte de la rede alimenticia de Dimetrodon, nutriéndose de pequeños reptiles como Captorhinus. Los Llechos Coloraos tamién fueron l'hábitat de dalgunos de los primeros grandes herbívoros terrestres como Edaphosaurus y Diadectes. Estos herbívoros que s'alimentaben de plantes terrestres nun dependíen del mediu acuáticu pa la so sobrevivencia. D'alcuerdu a Olson, l'análogu modernu más paecíu al ecosistema de Dimetrodon son los Everglades.[44]
La única especie de Dimetrodon topada fora del sureste d'Estaos Xuníos ye Dimetrodon teutonis d'Alemaña. Los sos restos fueron topaos na Formación Tambach nel sitiu fosilíferu Bromacker. Esti sitiu de tetrápodos del Permianu Inferior ye inusual pola escasez de sinápsidos depredadores de gran tamañu. La especie D. teutonis midía alredor de 1,7 metros, bien pequeña p'atrapar los grandes herbívoros diadéctidos que poblaben la zona. Ye más probable que s'alimentara d'invertebraos pequeños ya inseutos. Solo trés especímenes fósiles pueden corresponder a grandes depredadores; créese que pueden pertenecer varanópidos grandes o esfenacodontos pequeños y dambos ser depredadores potenciales de D. teutonis. En contraste a les tierres baxes pantanoses de los Llechos Coloraos de Texas, créese que los depósitos Bromacker teníen un ambiente montascoso ensin especies acuátiques. Ye posible que los grandes carnívoros rellumaren pola so ausencia por cuenta de la so dependencia de los anfibios acuáticos pa la so sobrevivencia.[16]
Especímenes nuevos
Sicasí delles especies de Dimetrodon podíen algamar gran tamañu, conócense munchos especímenes nuevos.[45] El paleontólogu Robert Bakker suxurió en 1982 que los adultos habitaben principalmente nes planicies inundables, mientres los mozos preferíen banzaos aisllaos y les márxenes de los llagos. El tipu d'ambiente deducir a partir del tipu de sedimentu nel cual fueron topaos los restos. Bakker pensó que los especímenes nuevos de Dimetrodon portar de manera similar a los mozos de los grandes reptiles modernos, que eviten el contautu colos adultos de la so especie. Nos reptiles actuales, los animales de toles edaes pueden competir polos mesmos recursos y los adultos pueden llegar a convertir a los mozos nes sos preses.[46] Sicasí, un estudiu posterior de Donald Brinkman pon en dulda l'afirmación de Bakker al demostrar que los especímenes topaos nos dos tipos d'hábitat representaben a especies distintes y non a miembros adultos y nuevos de la mesma especie.[47]
Especies
| Especie | Autoridá | Allugamientu | Estáu | Sinónimu | Imaxe |
|---|---|---|---|---|---|
| Dimetrodon angelensis | Olson, 1962 | Texas | Válidu |  | |
| Dimetrodon booneorum | Romer, 1937 | Texas | Válidu | ||
| Dimetrodon cruciger | Cope, 1878 | Texas | Sinónimu Edaphosaurus cruciger | ||
| Dimetrodon dollovianus | Case, 1907 | Texas | Válidu | Embolophorus dollovianus Cope, 1888 | |
| Dimetrodon giganhomogenes | Case, 1907 | Texas | Válidu |  | |
| Dimetrodon gigas | Cope, 1878 | Texas | Sinónimu de Dimetrodon grandis | Clepsydrops gigas Cope, 1878 | |
| Dimetrodon grandis | Romer y Price, 1940 | Oklahoma Texas | Válidu | Clepsydrops gigas Cope, 1878 Dimetrodon gigas Cope, 1878 Theropleura grandis Case, 1907 Bathyglyptus theodori Case, 1911 Dimetrodon maximus Romer 1936 |  |
| Dimetrodon incisivus | Cope, 1878 | Texas | Sinónimu de Dimetrodon limbatus | ||
| ?Dimetrodon kempae | Romer, 1937 | Texas | Posible nomen dubium | ||
| Dimetrodon limbatus | Romer y Price, 1940 | Oklahoma Texas | Válidu | Clepsydrops limbatus Cope, 1877 Dimetrodon incisivus Cope, 1878 Dimetrodon rectiformis Cope, 1878 Dimetrodon semiradicatus Cope, 1881 |  |
| Dimetrodon longiramus | Case, 1907 | Texas | Sinónimu de Secodontosaurus obtusidens | ||
| Dimetrodon loomisi | Romer, 1937 | Texas Oklahoma | Válidu |  | |
| Dimetrodon macrospondylus | Case, 1907 | Texas | Válidu | Clepsydrops macrospondylus Cope, 1884 Dimetrodon platycentrus Case, 1907 | |
| Dimetrodon milleri | Romer, 1937 | Texas | Válidu | | |
| Dimetrodon natalis | Romer, 1936 | Texas | Válidu | Clepsydrops natalis Cope, 1878 |  |
| Dimetrodon occidentalis | Berman, 1977 | Arizona Nuevu Méxicu Utah | Válidu | ||
| Dimetrodon platycentrus | Case, 1907 | Texas | Sinónimu de Dimetrodon macrospondylus | ||
| Dimetrodon rectiformis | Cope, 1878 | Texas | Sinónimu de Dimetrodon limbatus | ||
| Dimetrodon semiradicatus | Cope, 1881 | Texas | Sinónimu de Dimetrodon limbatus | ||
| Dimetrodon teutonis | Berman et al., 2001 | Alemaña | Válidu | ||
Ver tamién
- Llista de pelicosaurios
- Edaphosaurus
- Haptodus
- Sphenacodon
- Superpredador
Referencies
- ↑ «Dimetrodon». Paleobiology Database. Consultáu'l 23 d'agostu de 2012.
- 1 2 Angielczyk, K. D. (2009). «Dimetrodon is Not a Dinosaur: Using Tree Thinking to Understand the Ancient Relatives of Mammals and their Evolution». Evolution: Education and Outreach 2 (2): páxs. 257–271. doi:.
- ↑ Huttenlocker, A. K., and Y. Rega. 2012. The Paleobiology and Bone Microstructure of Pelycosauriangrade Synapsids. Pp. 90–119 in A. Chinsamy (ed.) Forerunners of Mammals: Radiation, Histology, Biology. Indiana University Press.
- ↑ Matthew, W.D. (1908). «Review of Case's 'Revision of the Pelycosauria of North America'». Science 27 (699): páxs. 816–818. https://books.google.com/books?id=5kAbAAAAYAAJ&printsec=frontcover#v=onepage&q&f=false.
- 1 2 Case, Y.C. (1907). Revision of the Pelycosauria of North America. Washington, D.C.: Carnegie Institution of Washington, páx. 1–176.
- 1 2 Cope, Y.D. (1878). «Descriptions of extinct Batrachia and Reptilia from the Permian formation of Texas». Proceedings of the American Philosophical Society 17 (101): páxs. 505–530.
- ↑ Cope, Y.D. (1877). «Descriptions of extinct vertebrata from the Permian and Triassic Formations of the United States». Proceedings of the American Philosophical Society 17 (100): páxs. 182–193.
- 1 2 3 Romer, A.S.; Price, L.I. (1940). «Review of the Pelycosauria». Geological Society of America Special Paper 28: páxs. 1–538.
- 1 2 3 Huttenlocker, A.K.; Rega, Y.; Sumida, S.S. (2010). «Comparative anatomy and osteohistology of hyperelongate neural spines in the sphenacodontids Sphenacodon and Dimetrodon (Amniota: Synapsida)». Journal of Morphology 271 (12): páxs. 1407–1421. doi:. http://onlinelibrary.wiley.com/doi/10.1002/jmor.10876/abstract.
- ↑ Vaughn, P.P. (1966). «Comparison of the Early Permian vertebrate faunes of the Four Corners region and north-central Texas». Los Angeles County Museum of Natural History Contributions in Science 105: páxs. 1–13.
- ↑ Vaughn, P.P. (1969). «Early Permian vertebrates from southern New Mexico and their paleozoogeographic significance». Los Angeles County Museum of Natural History Contributions in Science 166: páxs. 1–22.
- ↑ Olson, Y.C. (1975). «Permo-Carboniferous paleoecology and morphotypic series». American Zoologist 15 (2): páxs. 371–389.
- 1 2 Berman, D.S. (1977). «A new species of Dimetrodon (Reptilia, Pelycosauria) from a non-deltaic facies in the Lower Permian of north-central New Mexico». Journal of Paleontology 51 (1): páxs. 108–115.
- ↑ Madalena, K.; Sumida, S.; Zeigler, K.; Rega, Y. (2007). «A new record of the Early Permian pelycosaurian-grade synapsid Dimetrodon (Eupelycosauria: Sphenacodontidae) from the Lower Cutler Group (Early Permian) of Jemez Pueblu, north-central New Mexico». Journal of Vertebrate Paleontology 27 (3, Suppl.): páxs. 110A. doi:.
- ↑ Vaughn, P.P. (1973). New Mexico Geological Society: Vertebrates from the Cutler Group of Monument Valley and vicinity 24.
- 1 2 3 Berman, D.S.; Reisz, R.R.; Martens, T.; Henrici, A.C. (2001). «A new species of Dimetrodon (Synapsida: Sphenacodontidae) from the Lower Permian of Germany records first occurrence of genus outside of North America». Canadian Journal of Earth Sciences 38: páxs. 803–812. doi:. http://www.stuartsumida.com/BIOL680-09/BermanEtAl2001.pdf.
- 1 2 Palævos:The Vertebrates. «Dimetrodon» (inglés). Consultáu'l 26 d'ochobre de 2010.
- 1 2 Kenneth D. Angielczyk (2009). «Dimetrodon Is Not a Dinosaur: Using Tree Thinking to Understand the Ancient Relatives of Mammals and their Evolution». Evolution: Education and outreach 2 (2): p. 257-271. doi:. http://www.springerlink.com/content/w3o27142m0144433/fulltext.pdf.
- 1 2 Benson, R.J. (2012). «Interrelationships of basal synapsids: cranial and postcranial morphological partitions suggest different topologies». Journal of Systematic Paleontology in press. doi:. http://www.tandfonline.com/doi/abs/10.1080/14772019.2011.631042.
- ↑ Reisz, R.R., «Pelycosauria», en Sues, H.-D. (ed.), Pelycosauria, 17A, Gustav Fischer Verlag, pp. 1–102
- 1 2 Fröbisch, J.; Schoch, R.R.; Müller, J.; Schindler, T.; Schweiss, D. (2011). «A new basal sphenacodontid synapsid from the Late Carboniferous of the Saar-Nahe Basin, Germany». Acta Palaeontologica Polonica 56 (1): páxs. 113–120. doi:. http://app.pan.pl/archive/published/app56/app20100039.pdf.
- ↑ Olson, Y.C.; Beerbower, J.R. (1953). «The San Angelo Formation, Permian of Texas, and its vertebrates». The Journal of Geology 61 (5): páxs. 389–423.
- ↑ Olson, Y.C. (1955). «Parallelism in the evolution of the Permian reptilian faunes of the Old and New Worlds». Fieldiana 37 (13): páxs. 385–401. http://archive.org/stream/parallelisminevo3713olso#page/n5/mode/2up.
- ↑ Laurin, M.. «Synapsida: mammals and their extinct relatives». Tree of Life Web Project. Consultáu'l 24 d'agostu de 2012.
- 1 2 Herbert R. Barghusen (setiembre 1973). «The adductor jaw musculature of Dimetrodon (Reptilia, Pelycosauria)» (n'inglés). Journal of Paleontology 47 (5): páxs. 823-834. http://www.jstor.org/pss/1303062.
- ↑ «Exhibit Specimens: Dimetrodon». American Museum of Natural History. Archiváu dende l'orixinal, el 4 de xunetu de 2012. Consultáu'l 2 de xunetu de 2012.
- ↑ Baur, G.; Case, Y.C. (1899). «The history of the Pelycosauria, with a description of the genus Dimetrodon, Cope». Transactions of the American Philosophical Society 20 (1): páxs. 5–62.
- ↑ Kemp, T.S. (2006). «The origin and early radiation of the therapsid mammal-like reptiles: a palaeobiological hypothesis». Journal of Evolutionary Biology 19 (4): páxs. 1231–1247. doi:. PMID 16780524.
- ↑ Laurin, M.. «Autapomorphies of the main clades of synapsids». Tree of Life Web Project. Consultáu'l 24 d'agostu de 2012.
- ↑ Romer, A.S. (1927). «Notes on the Permo-Carboniferous reptile Dimetrodon.». The Journal of Geology 35 (8): páxs. 673–689.
- ↑ Rega, Y. A. (2012). «Healed Quebres in the Neural Spines of an Associated Skeleton of Dimetrodon: Implications for Dorsal Sail Morphology and Function». Fieldiana Life and Earth Sciences 5: páxs. 104. doi:.
- 1 2 3 Rega, Y.; Sumida, S.; Noriega, K.; Pell, C.; Lee, A. (2005). «Evidence-based paleopathology I: Ontogenetic and functional implications of dorsal sails in Dimetrodon.». Journal of Vertebrate Paleontology 25 (S3): páxs. 103A.
- ↑ Sumida, S.; Rega, Y.; Noriega, K. (2005). «Evidence-based paleopathology II: Impact on phylogenetic analysis of the genus Dimetrodon.». Journal of Vertebrate Paleontology 25 (S3): páxs. 120A.
- ↑ Rega, Y.A.; Noriega, K.; Sumida, S.; Lee, A. (2004). «Histological analysis of traumatic injury to multiple neural spines of an associated skeleton of Dimetrodon: Implications for healing response, dorsal sail morphology and age-at-death in a Lower Permian synapsid». Integrated Comparative Biology 44: páxs. 628.
- ↑ Rega, Y. A. (2012). «Healed Quebres in the Neural Spines of an Associated Skeleton of Dimetrodon:Implications for Dorsal Sail Morphology and Function». Fieldiana Life and Earth Sciences 5: páxs. 104. doi:.
- 1 2 Florides, G.A.; Kalogirou, S.A.; Tassou, S.A.; Wrobel, L. (2001). «Natural environment and thermal behaviour of Dimetrodon limbatus.». Journal of Thermal Biology 26 (1): páxs. 15–20. doi:. http://www.sciencedirect.com/science/article/pii/S030645650000019X.
- ↑ Bramwell, C.D.; Fellgett, P.B. (1973). «Thermal regulation in sail lizards». Nature 242 (5394): páxs. 203–205. doi:. http://www.nature.com/nature/journal/v242/n5394/pdf/242203a0.pdf.
- 1 2 Haack, S.C. (1986). «A thermal model of the sailback pelycosaur». Paleobiology 12 (4): páxs. 450–458.
- ↑ Florides, G.A.; Wrobel, L.C.; Kalogirou, S.A.; Tassou, S.A. (1999). «A thermal model for reptiles and pelycosaurs». Journal of Thermal Biology 24 (1): páxs. 1–13. doi:. http://www.sciencedirect.com/science/article/pii/S0306456598000321.
- ↑ Turner, J.S.; Tracy, C.R. (1986). «Body size, homeothermy and the control of heat exchange in mammal-like reptiles», The Ecology and Biology of Mammal-Like Reptiles. Washington, D.C.: Smithsonian Institution Press, páx. 185–194.
- 1 2 Tomkins, J.L.; LeBas, N.R.; Witton, M.P.; Martill, D.M.; Humphries, S. (2010). «el%20Am%20Nat%202010.pdf Positive allometry and the prehistory of sexual selection». The American Naturalist 176 (2): páxs. 141–148. doi:. PMID 20565262. http://www.alternativetactics.org/PDF/Tomkins%20et%20a el%20Am%20Nat%202010.pdf.
- ↑ Lucas, S.G.; Spielmann, J.A.; Rinehart, L.F.; Martens, T. (2009). New Mexico Geological Society: Dimetrodon (Amniota: Synapsida: Sphenacodontidae) from the Lower Permian Abo Formation, Socorru County, New Mexico 60.
- ↑ Sullivan, C.; Reisz, R.R.; May, W.J. (2000). «Large dissorophoid skeletal elements from the Lower Permian Richards Spur fissures, Oklahoma, and their paleoecological implications». Journal of Vertebrate Paleontology 20 (3): páxs. 456–461. doi:.
- 1 2 Olson, Y.C. (1966). «Community evolution and the origin of mammals». Ecology 47 (2): páxs. 291–302.
- ↑ Sternberg, C.W. (1942). «The skeleton of an immature pelycosaur, Dimetrodon cf. grandis, from the Permian of Texas». Journal of Paleontology 16 (4): páxs. 485–486.
- ↑ Bakker, R. T. (1982). «Juvenile-Adult Habitat Shift in Permian Fossil Reptiles and Amphibians». Science 217 (4554): páxs. 53–55. doi:. PMID 17739981.
- ↑ Brinkman, D. (1988). «Size-independent criteria for estimating relative age in Ophiacodon and Dimetrodon (Reptilia, Pelycosauria) from the Admiral and lower Belle Plains formations of west-central Texas». Journal of Vertebrate Paleontology 8 (2): páxs. 172–180.
Bibliografía
- Berman, D.S., Reisz, R.R., Martens, T., & Henrici, A.C. 2001. A new species of Dimetrodon (Synapsida: Sphenacodontidae) from the Lower Permian of Germany records first occurrence of genus outside of North America. Can. J. Earth Sci., 38(5): 803–812
Enllaces esternos
| Wikispecies tien un artículu sobre Dimetrodon. |
- http://www.juntadeandalucia.es/averroes/ies_torre_d'el_aguila/DINO/dimetrodon.htm
- http://www.enchantedlearning.com/subjects/dinosaurs/dinos/Dimetrodon.shtml (inglés)
- http://www.prehistory.com/dimetrod.htm
| Control d'autoridaes |
|
|---|
 Datos: Q130869
Datos: Q130869 Multimedia: Dimetrodon
Multimedia: Dimetrodon Especies: Dimetrodon
Especies: Dimetrodon