
Los cromatóforos son célules con pigmentos nel so interior que reflexen la lluz. Pueden atopase en diversos seres vivos como los anfibios, los peces, ciertos crustáceos y dalgunos cefalópodos. Son los principales responsables de la coloración de la piel, del color de los güeyos nos animales ectotermos y de la formación de la cresta neural a lo llargo del desenvolvimientu embrionariu. Los cromatóforos yá maduros pueden estremase en distintes clases según el color que reflexen so una lluz blanco: cianóforos (azul), eritróforos (colloráu), iridóforos (iridiscente), leucóforos (blancu), melanóforos (negru/marrón) y xantóforos (mariellu). El términu tamién puede faer referencia a les visícules coloriaes asociaes a la membrana de ciertes bacteries fotosintétiques.
Delles especies pueden camudar de color rápido per mediu de mecanismos que camuden un pigmentu por otru y reorienten la superficie reflectora de los cromatóforos. Esti procesu, de cutiu utilizáu como mecanismu de camuflaje, ye llamáu mimetismu o cambéu fisiolóxicu de color. Dellos cefalópodos como los pulpos, presenten unos órganos cromatóforos realmente complexos, controlaos por músculos, ente que ciertos vertebraos, como'l camaleón, producen efeutos similares mediaos nesti casu per víes de señalización celular. Estes señales pueden ser hormones o neurotransmisores y pueden ser inducíes por cambeos nel humor, la temperatura, el estrés o cambeos notables del ambiente a nivel llocal.
A diferencia de los animales ectotermos, los mamíferos y los páxaros presenten namái una clase de célules del tipu de los cromatóforos: los melanocitos. L'equivalente nos animales ectotermos, los melanóforos, son estudiaos anguaño nos llaboratorios d'investigación col fin d'ayudar na comprensión de ciertes enfermedaes humanes y utilizaos como ferramienta nel descubrimientu o diseñu de nuevos fármacos.
Clasificación
Les primeres célules pigmentadas descrites pertenecíen a invertebraos y fueron denominaes cromóforos nuna revista científica italiana nel añu 1819.[1] El términu cromatóforu acuñóse más tarde pa denominar a les célules que conteníen pigmentos provenientes de la cresta neural de los vertebraos ectotermos y de los cefalópodos. La pallabra cromatóforu provién del griegu y puede estremase en dos términos:
- khrōma (χρωμα), que'l so significáu ye color.
- phoros (φορος), que'l so significáu ye que lleva.
Otra manera, la pallabra cromatocito (cito o κυτε del griegu, que'l so significáu ye célula) foi acuñada pa denominar a les célules responsables del color atopaes en páxaros y mamíferos, anque namái atopóse unu d'estos tipos celulares, el melanocito.
Sicasí, la estructura y la coloración de los cromatóforos nun foi desentrañada hasta la década de los 60, momentu a partir del cual pudo desenvolvese un sistema de clasificación basáu na apariencia de dichos cromatóforos. Esta clasificación caltúvose hasta los nuesos díes anque los últimos estudios al respeutu punxeron de manifiestu dellos aspeutos bioquímicos de los pigmentos que podríen ser más útiles a la d'entender la so función na célula.[2]
Según la producción de color, los cromatóforos pueden estremase en dos clases:
- Biocromos: presenten pigmentos verdaderos, como pueden ser los carotenoides y les pteridines. Estos pigmentos absuerben selectivamente determinaes franxes del espectru de lluz visible, que se compón de lluz blanco, emitiendo asina aquelles llonxitúes d'onda que nun absuerben, y que son les que lleguen al güeyu del observador.
- Esquemocromos: tamién conocíos como colores estructurales, xeneren la so coloración bien por reflexón de ciertos llonxitúes d'onda (colores) de lluz visible y emisión d'otres, bien xenerando ondes de lluz qu'interfieren dientro de la estructura o bien esvalixando la lluz que-yos llega.
Anque tolos cromatóforos contienen pigmentos o estructures reflectantes (sacante cuando fueron la resultancia d'una mutación xenética como nel casu del albinismu), non toles célules que contienen pigmentos son cromatóforos. Por casu, el grupu hemo ye un biocromo responsable del color coloráu de la sangre. Atópase principalmente nos eritrocitos, que son xeneraos nel migollu oseu constantemente a lo llargo de la vida d'un organismu y non namái mientres el desenvolvimientu embrionariu. Por ello, los eritrocitos anque tengan pigmentos, al nun provenir de les célules de la cresta neural, nun son clasificaos como cromatóforos.

Xantóforos y eritróforos
Esti tipu de cromatóforos contién, o bien una gran cantidá de pigmentos mariellos del tipu de les pteridinas y son denominaos xantóforos, o bien una gran cantidá de pigmentos colloraos/naranxes del tipu de los carotenoides, denominándose entós eritróforos.[3] Sía que non, púdose reparar que les visícules que contienen estos pigmentos, les pteridinas y los carotenoides, pueden llegar a atopase na mesma célula, lo qu'amplía en gran midida l'abanicu de colores que pueden reparase, dependiendo de la rellación ente pigmentos coloraos y mariellos.[4] Por ello, la distinción fecha ente estos cromatóforos realmente ye arbitraria. La capacidá de xenerar pteridinas dende guanosín trifosfato ye una carauterística común a la mayoría de los cromatóforos, pero los xantóforos paecen tener rutes bioquímiques alternatives que dan llugar a un escesu de pigmentos mariellos acumulaos. Otra manera, los carotenoides son metabolizados a partir de los alimentos inxeríos na dieta y tresportaos a los eritróforos. Esto foi demostráu per primer vegada con un senciellu esperimentu. Criáronse les típiques xaronques verdes sometiéndoles a una dieta onde s'esaniciaron dafechu los carotenos. L'ausencia de carotenos dio llugar a una ausencia de pigmentos coloraos/naranxes nos eritróforos, y esto traducióse en que les xaronques paecíen ser de color azul en llugar de verdes.[5]
Iridóforos y leucóforos
Los iridóforos, tamién denominaos guanóforos, son pigmentos celulares que reflexen la lluz usando llámines d'esquemocromos cristalinos sintetizaos a partir de guanina.[6] Cuando estes llámines son allumaes xeneren colores iridiscentes por cuenta de la difracción que sufren los rayos de lluz al travesar la pila de llámines. La orientación de los esquemocromos va determinar la naturaleza del color reparáu.[7] Cuando los iridóforos utilicen biocromos como filtros coloriaos xeneren un efeutu ópticu conocíu como efeutu Tyndall o dispersión de Rayleigh, lo cual da llugar a la producción de colores azules brilloses y verdes.[8]
Esiste otru cromatóforu rellacionáu, los leucóforos, que puede atopase en ciertes especies de peces. Al igual que los iridóforos, utilicen purines cristalines pa reflexar la lluz, xenerando asina'l típicu rellumu de color blancu que puede apreciase en dellos pexes. La distinción ente iridóforos y leucóforos en peces nun ye siempres evidente, al igual que pasa colos xantóforos y los eritróforos, pero xeneralmente, los iridóforos producen colores iridiscentes o metálicu ente que los leucóforos xeneren reflexos de diversos tonos de blancu.[8]
Melanóforos
Los melanóforos contienen eumelanina, un tipu de melanina de color negru o marrón escuru por cuenta de la so gran capacidá p'absorber la mayoría de los llonxitúes d'onda de la lluz. Atópase nel interior d'unes visícules denominaes melanosomes, que se distribuyen por tol citoplasma celular. La eumelanina sintetizar a partir de la tirosina al traviés d'una serie de reacciones de catálisis química. Presenta una estructura complexa basada n'unidaes de dihidroxindol y dihidroxindol-2-carboxilato con dellos aniellos pirrólicos.[9] La enzima clave na síntesis de melanina ye la tirosinasa. Si esta enzima atópase ausente, la melanina nun puede sintetizase y esto desagua en diversos tipos d'albinismu.
En delles especies d'anfibios pueden apreciase unos pigmentos distintos xunto a la eumelanina, como por casu, un nuevu pigmentu de color acoloratáu que foi identificáu nos melanóforos de filomedusina de ciertes xaronques.[10] Darréu, esti pigmentu foi identificáu como pterorodina, un dímero de pteridina que s'atropa alredor de la eumelanina. Anque probablemente haya más subtipos de pigmentos nos melanóforos, hasta agora, na mayoría de los melanóforos estudiaos conteníen puramente eumelanina.
Los seres humanos tienen tan solo una clase de célula pigmentada, l'equivalente mamíferu de los melanóforos, que s'encarga de xenerar el color de la piel, el pelo y los güeyos. Por ello, y polo fácil que ye estudiar los debíu al eleváu númberu de colores distintos que presenten, los melanóforos son, con muncha diferencia, los cromatóforos más llargamente estudiaos. Sicasí, esisten diferencies ente la bioloxía de los melanóforos y la de los melanocitos. Amás de la eumelanina, los melanocitos pueden sintetizar pigmentos mariellos/verdes denominaos feomelaninas.

Cianóforos
En 1995 demostróse que los colores azul brillosu de dellos tipos de pexe mandarín (Siniperca chuatsi) nun yeren xeneraos por esquemocromos, sinón por un biocromo cian d'estructura química desconocida.[8] Esti pigmentu, atopáu nel interior de visícules en siquier dos especies de pexe de la familia Callionymidae, ye bien pocu común nel reinu animal, razón pola cual la mayoría de los pigmentos azules estudiaos hasta'l momentu son esquemocromáticos. Por ello, estos nuevos pigmentos propunxéronse como un nuevu tipu de cromatóforo, los cianóforos. Anque paecen tar pocu estendíos, ye posible qu'haya cianóforos (según otres clases de cromatóforos pocu comunes o desconocíos) n'otres especies de pexes y anfibios. Por casu, los cromatóforos de colores brillosos que se repararon na xaronca venenosa de flecha (familia Dendrobatidae) y na xaronca de cristal (familia Centrolenidae), que los sos pigmentos son desconocíos.[11]
Translocación de pigmentos
Munches especies tienen la capacidá de translocar los pigmentos nel interior de los cromatóforos, lo que da llugar a un aparente cambéu de color nel animal. Esti procesu, conocíu como cambéu fisiolóxicu de color, ta llargamente estudiáu nos melanóforos, por cuenta de que la melanina ye'l pigmentu más escuru y por ello'l más visible. Na mayoría de les especies que tienen una dermis relativamente delgada, los melanóforos de la dermis tienden a ser planos y a cubrir una gran superficie. Sicasí, nos animales que la so dermis ye gruesa, como los reptiles adultos, los melanóforos de la dermis suelen formar unidaes tridimensionales con otros melanóforos. Estes unidaes dérmiques de cromatóforos (UDC) componer d'una primer capa cimera de xantóforos o eritróforos, siguida d'una capa de iridóforos y finalmente una especie de capa de melanóforos.[12]
Dambos tipos de melanóforos son importantes nel procesu del cambéu fisiolóxicu de color. Los melanóforos planos de la dermis suélense superponer a otros cromatóforos, polo que, cuando los pigmentos espublicen al traviés de la célula, la piel apaez escura. Otra manera, cuando los pigmentos concentrar nel centru de la célula, queden espuestos a la lluz los pigmentos de los cromatóforos que s'atopen nes capes inferiores, lo que produz que la piel tornar n'otres tonalidaes. De forma similar, en amestándose la melanina en UDCs, la piel repara verde debíu al filtru qu'exercen los xantóforos (mariellos) de la lluz tremao que llega dende la capa de los iridóforos. Cuando se produz l'espardimientu de la melanina, la lluz nun s'esvalixa tantu y la piel apaez escura. Al igual qu'otros cromatóforos biocromáticos que son capaces de translocar pigmentos, los animales con múltiples tipos de cromatóforos pueden xenerar un espectacular abanicu de colores na so piel, aprovechando l'efeutu divisional.[13],[14]

El control y los mecanismos implicaos na translocación rápida de pigmentos fueron llargamente estudiaos nun determináu númberu d'especies, ente los que cabo destacar a los anfibios y a los pexes teleósteos.[15],[8] Demostróse qu'esti procesu puede tar reguláu por procesos hormona-yos, neuronales o por dambos. Describióse que ciertos neurotransmisores como la noradrenalina, tán rellacionaos cola translocación de pigmentos per mediu de receptores na superficie de los melanóforos.[16] Ente les principales hormones arreyaes na regulación d'esti procesu atopen les melanocortines, la melatonina y la hormona concentrante de melanina (MCH), que son producíes principalmente na glándula pituitaria, na glándula pineal y nel hipotálamu, respeutivamente. Estes hormones tamién podríen ser xeneraes de manera paracrino poles célules de la piel. Tamién se reparó que, na superficie de los melanóforos, les hormones activen específicamente al receptor acopláu a proteínes G, que transduce la señal al interior de la célula. Los efeutos que producen estes hormones son diversos. Les melanocortinas dan llugar a la dispersión de los pigmentos, ente que la melatonina y la MHC dan llugar al agregamientu de los mesmos.[17]
Fueron identificaos ensame de receptores de melanocortina, de MHC y de melatonina en ciertes especies de pexes[18] y xaronques,[19] incluyendo un homólogu de MC1R,[20] un receptor de melanocortina conocíu por regular el procesu de coloración de la piel y del pelo nos humanos.[21] De la mesma, demostróse que, nel interior celular, l'AMPc ye un importante segundu mensaxeru de la translocación pigmentaria. Al traviés d'un mecanismu entá non entendíu dafechu, el AMPc actúa sobre otres enzimes tales como la proteinasa K, col fin d'activar motores moleculares que tresporten les visícules con pigmentos a lo llargo de los microtúbulos y los microfilamentos.[22],[23],[24]
Adautación al fondu
La mayoría de los pexes, reptiles y anfibios esperimenten un cambéu fisiolóxicu de color llindáu en respuesta a un cambéu nel ambiente. Esti tipu de camuflaje, conocíu como adautación al fondu, manifiéstase de normal como un llixeru escurecimientu o aclaramiento de la tonalidá de la piel, que'l so fin ye mimetizar el color y la tonalidá del mediu ambiente nel que s'atope. Demostróse que'l procesu d'adautación al fondu ye dependiente de la visión (paez que ye necesariu que l'animal vea l'ambiente nel que s'atopa pa ser capaz d'afaer el color de la so piel a dichu entorno),[25] y que la translocación de melanina nos melanóforos ye'l factor principal nel cambéu de color.[17] Dellos animales, como'l camaleón o los anoles, desenvolvieron una efeutiva respuesta d'adautación al fondu, capaz de xenerar un gran númberu de colores en curtios espacios de tiempu. Afixeron la capacidá pa camudar de color por que pueda responder a temperatura, cambeos d'humor, niveles de estrés, etc. y non a cencielles a cambeos ambientales.

Orixe embrionariu y desenvolvimientu
Mientres el desenvolvimientu embrionariu de los vertebraos, los cromatóforos pertenecen a unu de los tipos celulares xeneraos na cresta neural, un par de franxes de célules incipientes nos márxenes del tubu neural. Estes célules presenten la capacidá de migrar llargues distancies, lo que dexa que los cromatóforos lleguen y establézanse en distintos órganos del cuerpu, ente los que cabo destacar la piel, los güeyos, les oreyes y el celebru. N'abandonando la cresta neural en folaes, los cromatóforos tomen dos rutes posibles: o bien la ruta dorsolateral al traviés de la dermis, llegando al ectodermo al traviés de pequeños furacos na llámina basal, o bien la ruta ventromedial ente les somitas y el tubu neural. Una esceición a esto son los melanóforos del epiteliu pigmentado de la retina del güeyu, que nun deriven de la cresta neural, sinón d'una invaginación del tubu neural que xenera la copa óptica y esta de la mesma la retina.
Cuándo y cómo les célules precursores multipotentes de cromatóforos (denominaes cromatoblastos) dan llugar a los diversos subtipos de célules fíes ye oxetu d'investigación anguaño. Por casu, sábese que nos embriones de pez cebra, 3 díes dempués de la fertilización, caúna de les clases celulares atopaes nel pexe adultu — melanóforos, xantóforos y iridóforos — yá tán presentes. Estudios realizaos con mutantes d'esti pexe demostraron que ciertos factores de trescripción como kit, los xenes SOX y mitf son claves nel control de la diferenciación de los cromatóforos.[26] Cuando estes proteínes atópense ausentes los cromatóforos pueden tar parcial o totalmente ausentes, lo que da llugar a desordes leucísticos.
Aplicaciones práutiques
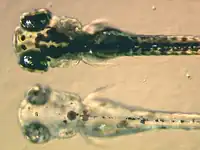
Amás de la investigación básica llevada a cabu pa llograr una meyor comprensión de los cromatóforos, buscáronse diverses utilidaes a estes célules. Por casu, los bárabos de los pexes zebra son utilizaes na actualidá pa estudiar cómo los cromatóforos entámense y comuniquen col fin de xenerar de forma desaxeradamente precisa'l típicu patrón de rayes horizontales que puede reparase nos individuos adultos.[27] Esto foi tremendamente útil a la de buscar un sistema modelu na creación de patrones col que trabayar nel campu de la bioloxía evolutiva del desenvolvimientu. La bioloxía de los cromatóforos tamién foi utilizada como modelu de ciertes enfermedaes humanes, como'l casu del melanoma, o de delles condiciones xenétiques, como'l albinismu. Coles mesmes, demostróse que les variaciones de pigmentación más clares, tantu nos seres humanos como na variedá "dorada" de pez cebra, deber a un menor númberu, tamañu y densidá de los melanosomes, los orgánulos pigmentados de los melanocitos. Tal carauterística ta acomuñada a un xen que codifica pa un intercambiador de cationes alcontráu nuna membrana intracelular denomináu Slc24a5.[28] El xen ortólogo n'humanos presenta dos alelos principales, que difieren nun solu nucleótido que determina un cambéu de alanina a treonina na posición 111 de la proteína. L'apanfilo ancestral, que lleva la alanina, foi topáu nel 93 a 100% de les muestres d'africanos, asiáticos del este y poblaciones orixinaries d'América. Sicasí, l'apanfilo que lleva la treonina, aniciáu fai 6.000 a 12.000 años tras y acomuñáu a pigmentación más clara, ta presente nel 98,7 a 100% de les muestres tomaes de poblaciones europees.[29] Poro, les variaciones nesti xen, allugáu nel cromosoma 15 del xenoma humanu, taríen esplicando les diferencies na coloración de la piel na nuesa especie[30]
Los cromatóforos son tamién usaos como biomarcadores de ceguera n'especies ectotermas, yá que, cuando un animal con un determináu defectu visual ye incapaz d'afaer el color de la so piel al ambiente, puede deducise que presenta un problema de visión.[25] Créese que los receptores homólogos n'humanos, que medien la translocación pigmentaria nos melanóforos, tán arreyaos en procesos tales como la perda de mambís y el morenura, lo que los convierte en posibles dianes a la de buscar fármacos rellacionaos.[31] Por ello, les compañíes farmacéutiques desenvolvieron ensayos biolóxicos pa la rápida identificación de posibles compuestos bioactivos, usando pa ello los melanóforos de la xaronca con garra africana (Xenopus laevis).[32] Otros científicos desenvolvieron téuniques col fin d'utilizar melanóforos como biosensores,[33] y pa la rápida detección d'enfermedaes (basáu nel descubrimientu de que la toxina producida pola bacteria Bordetella pertussis bloquia l'agregamientu de pigmentos nos melanóforos de los pexes).[34] A lo último, cabo destacar les posibles aplicaciones de los cromatóforos nel campu militar, onde podríen utilizase col fin de consiguir rápidos cambeos de color, y principalmente, como una forma de camuflaje activu.[35]
Cromatóforos de cefalópodos

Los cefalópodos de la subclase coleoidea presenten complexos órganos multicelulares qu'utilicen pa camudar de color rápido, como puede reparase nos calamares brillosos, les sepies y los pulpos. Cada unidá cromatófora ta compuesta por una única célula cromatófora y numberosos músculos, nervios, célules gliales y célules de la vaina.[36] Nel interior de la célula cromatófora, los gránulos de pigmentos atópense zarraos nun sacu elásticu, denomináu cytoelastic sacculus. Pa camudar de color, l'individuu deforma el tamañu o la forma del sacculus per mediu de contraiciones musculares, llogrando asina variar l'estáu de translucidez, reflexón o opacidá de los pigmentos. Esti mecanismu difier del utilizáu polos pexes, anfibios y reptiles, onde lo qu'asocede ye una translocación de pigmentos nel interior de la célula. Sicasí, l'efeutu que se repara a la fin ye bien similar.
Los pulpos presenten los cromatóforos nun complexu, capaz d'emitir distintes llonxitúes d'onda cromátiques, lo que da llugar a esquemes de colores que camuden rápido. Créese que los nervios que controlen a los cromatóforos asítiase nel celebru, nun orde similar a los cromatóforos que controlen. Esto quier dicir que'l patrón de cambéu de color ta d'acordies col patrón d'activación neuronal. Esto podría esplicar por qué, cuando les neurones son activaes una tres otra, el cambéu de color producir en forma d'ondes.[37] Al igual que los camaleones, los cefalópodos utilicen el cambéu fisiolóxicu de color como mediu d'interacción social. Amás, atopar ente los que tienen una mayor habilidá na adautación al fondu, presentando la capacidá d'asemeyar el so aspeutu tanto a los color como a la testura del ambiente nel que s'atopen de forma escepcionalmente precisa.
Cromatóforos en bacteries
Los cromatóforos tamién pueden atopase nes membranes de bacteries fototróficas, que los utilicen pa llevar a cabu la fotosíntesis y compónense de pigmentos de bacterioclorofila y carotenoides.[38] Nes bacteries púrpura como Rhodospirillum rubrum, les proteínes colectoras de lluz atopar de forma intrínseca nes membranes de los cromatóforos. Sicasí, les bacteries verdes del azufre desenvolvieron un complexu-antena especializáu, denomináu clorosoma.[39]
Ver tamién
- Camuflaje
- Mimetismu
Notes
- ↑ Sangiovanni G. Descrizione di un particolare sistema di organi cromoforo espansivo-dermoideo y dei fenomeni che esso produz, scoperto nei molluschi cefaloso. G. Enciclopedico Napoli. 1819; 9:1–13.
- ↑ Bagnara JT. Cytology and cytophysiology of non-melanophore pigment cells. Int Rev Cytol. 1966; 20:173–205. PMID 5337298
- ↑ Bagnara JT. Cytology and cytophysiology of non-melanophore pigment cells. Int Rev Cytol. 1966; 20:173–205. PMID 5337298
- ↑ Matsumoto J. Studies on fine structure and cytochemical properties of erythrophores in swordtail, Xiphophorus helleri. J Cell Biol. 1965; 27:493–504. PMID 5885426
- ↑ Bagnara JT. Comparative Anatomy and Physiology of Pigment Cells in Nonmammalian Tissues in The Pigmentary System: Physiology and Pathophysiology, Oxford University Press, 1998. ISBN 0-19-509861-7
- ↑ Taylor JD. The effects of intermedin on the ultrastructure of amphibian iridophores. Xen Comp Endocrinol. 1969; 12:405-16. PMID 5769930
- ↑ Morrison RL. A transmission electron microscopic (TEM) method for determining structural colors reflected by lizard iridophores. Pigment Cell Res. 1995; 8:28–36. PMID 7792252
- 1 2 3 4 Fujii R. The regulation of motile activity in fish chromatophores. Pigment Cell Res. 2000; 13:300-19. PMID 11041206
- ↑ Ito S & Wakamatsu K. Quantitative analysis of eumelanin and pheomelanin in humans, mice, and other animals: a comparative review. Pigment Cell Res. 2003; 16:523-31. PMID 12950732
- ↑ Bagnara JT et al. Color changes, unusual melanosomes, and a new pigment from leaf frogs. Science. 1973; 182:1034–5. PMID 4748673
- ↑ Schwalm PA et al. Infrared reflectance in leaf-sitting neotropical frogs. Science. 1977; 196:1225–7. PMID 860137
- ↑ Bagnara JT et al. The dermal chromatophore unit. J Cell Biol. 1968; 38:67–79. PMID 5691979 Full text — PDF.
- ↑ Palazzo RE et al. Rearrangements of pterinosomes and cytoskeleton accompanying pigment dispersion in goldfish xanthophores. Cell Motil Cytoskeleton. 1989; 13:9–20. PMID 2543509
- ↑ Porres MG et al. Corazonin promotes tegumentary pigment migration in the crayfish Procambarus clarkii. Peptides. 2003; 24:1581–9. PMID 14706537
- ↑ Deacon SW et al. Dynactin is required for bidirectional organelle transport. J Cell Biol. 2003; 160:297-301. PMID 12551954 Full text
- ↑ Aspengren S et al. Noradrenaline- and melatonin-mediated regulation of pigment aggregation in fish melanophores. Pigment Cell Res. 2003; 16:59–64. PMID 12519126
- 1 2 Logan DW et al. Regulation of pigmentation in zebrafish melanophores. Pigment Cell Res. 2006; 19:206-13. PMID 16704454
- ↑ Logan DW et al. Sequence characterization of teleost fish melanocortin receptors. Ann N Y Acad Sci. 2003; 994:319-30. PMID 12851332
- ↑ Sugden D et al. Melatonin, melatonin receptors and melanophores: a moving story. Pigment Cell Res. 2004; 17:454-60. PMID 15357831
- ↑ Logan DW et al. The structure and evolution of the melanocortin and MCH receptors in fish and mammals. Genomics. 2003; 81:184-91. PMID 12620396
- ↑ Valverde P et al. Variants of the melanocyte-stimulating hormone receptor gene are associated with red hair and fair skin in humans. Nat Genet. 1995; 11:328-30. PMID 7581459
- ↑ Snider J et al. Intracellular actin-based transport: how far you go depends on how often you switch. Proc Natl Acad Sci USA. 2004; 101:13204-9. PMID 15331778 Full text
- ↑ Rodionov VI et al. Functional coordination of microtubule-based and actin-based motility in melanophores. Curr Biol. 1998; 8:165-8. PMID 9443917 Full text
- ↑ Rodionov VI et al. Protein kinase A, which regulates intracellular transport, forms complexes with molecular motors on organelles. Curr Biol. 2002; 14:1877–81. PMID 15498498 Full text
- 1 2 Neuhauss SC. Behavioral genetic approaches to visual system development and function in zebrafish. J Neurobiol. 2003; 54:148-60. PMID 12486702. Full text — PDF.
- ↑ Kelsh RN et al. Genetic analysis of melanophore development in zebrafish embryos.] Dev Biol. 2000; 225:277-93. PMID 10985850
- ↑ Kelsh RN. Genetics and evolution of pigment patterns in fish. Pigment Cell Res. 2004; 17:326-36. PMID 15250934
- ↑ Lamason RL et al. SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans. Science. 2005; 310:1782–6. PMID 16357253
- ↑ Peen State University: fish gene sheds light on human skin color variation. Archiváu el 21 de xunetu de 2010 na Wayback Machine.
- ↑ Gibbons A (2007). «American Association of Physical Anthropologists meeting. European skin turned pale only recently, gene suggests». Science 316 (5823): páxs. 364. doi:. PMID 17446367. http://img46.imageshack.us/img46/4784/eurospaleonlyrecentlypu0.jpg.
- ↑ Logan DW et al. The structure and evolution of the melanocortin and MCH receptors in fish and mammals. Genomics. 2003; 81:184-91. PMID 12620396.
- ↑ Jayawickreme CK et al. Use of a cell-based, lawn format assay to rapidly screen a 442,368 bead-based peptide library. J Pharmacol Toxicol Methods. 1999; 42:189-97. PMID 11033434
- ↑ Andersson TP et al. Frog melanophores cultured on fluorescent microbeads: biomimic-based biosensing. Biosens Bioelectron. 2005; 21:111-20. PMID 15967358
- ↑ Karlsson JO et al. The melanophore aggregating response of isolated fish scales: a very rapid and sensitive diagnosis of whooping cough. FEMS Microbiol Lett. 1991; 66:169-75. PMID 1936946
- ↑ Lee I. Nanotubes for noisy signal processing: Adaptive Camouflage PhD Thesis. 2005; University of Southern California. Retrieved June 2006 — PDF (799 KiB).
- ↑ Cloney RA. & Florey Y. Ultrastructure of cephalopod chromatophore organs. Z Zellforsch Mikrosk Anat. 1968; 89:250–280. PMID 5700268
- ↑ Demski LS. Chromatophore systems in teleosts and cephalopods: a levels oriented analysis of convergent systems. Brain Behav Evol. 1992; 40:141-56. PMID 1422807
- ↑ Salton MR. Bacterial membrane proteins. Microbiol Sci. 1987; 4:100-5. PMID 3153178
- ↑ Frigaard NU. & Bryant DA. Seeing green bacteria in a new light: genomics-enabled studies of the photosynthetic apparatus in green sulfur bacteria and filamentous anoxygenic phototrophic bacteria. Arch Microbiol. 2004; 182:265-75. PMID 15340781
{kind=link}
Enllaces esternos
- Videu de l'adautación al fondu d'un pulpu
- Videu del patrón de cromatóforu d'un calamar
- Proyeutu Web Árbol de la Vida: cromatóforos de cefalópodos
| Control d'autoridaes |
|
|---|
 Datos: Q504699
Datos: Q504699 Multimedia: Chromatophores
Multimedia: Chromatophores