Plantía:Ficha de familia de proteínes L'actina ye una familia de proteínes globulares que formen los microfilamentos, unu de los trés componentes fundamentales del citoesqueleto de les célules de los organismos eucariotes (tamién denominaos eucariontes). Puede atopase como monómero en forma llibre, denominada actina G, o como parte de polímeros lliniales denominaos microfilamentos o actina F, que son esenciales pa funciones celulares tan importantes como la movilidá y la contraición de la célula mientres la división celular.
De la importancia capital de la actina da cuenta'l fechu de que nel conteníu proteico d'una célula supón siempres un eleváu porcentaxe y que'l so secuencia ta bien caltenida, esto ye, que camudó bien pocu a lo llargo de la evolución.[1][2] Por dambes razones puede dicise que la so estructura foi optimizada. Sobre ésta puédense destacar dos traces peculiares: ye una enzima qu'hidroliza ATP, la "moneda universal de la enerxía" de los procesos biolóxicos, faciéndolo bien amodo. Pero coles mesmes precisa d'esa molécula pa caltener la so integridá estructural. Adquier la so forma eficaz nun procesu de plegamiento cuasi dedicáu. Amás ye la qu'establez más interaiciones con otres proteínes de cuantes conócense, lo que-y dexa desempeñar les más variaes funciones qu'algamen a cuasi tolos aspeutos de la vida celular. La miosina ye un exemplu de proteína que xune actina. Otru exemplu ye la vilina, que puede enxareyar la actina en fexes o bien cortar los filamentos d'actina, dependiendo de la concentración de catión calciu na so redolada.[3]
Formando microfilamentos nun procesu dinámicu apurre un andamiaxe que dota a la célula d'una forma con posibilidá de remocicase rápido en respuesta a la so redolada o a señales del organismu, por casu, aumentando la superficie celular pa l'absorción o apurriendo soporte a l'adhesión de les célules pa formar texíos. Sobre esti andamiaxe pueden fondiase otres enzimes, orgánulos como'l cilio, dirixir la deformación de la membrana celular esterna que dexa la ingestión celular o la citocinesis. Tamién puede producir movimientu, bien por ella mesma o ayudada de motores moleculares. D'esa manera contribúi a procesos como'l tresporte intracelular de visícules y orgánulos y la contraición muscular, o la migración celular, importante nel desarrollu embrionariu, arreglu de firíes o invasividad del cáncer. L'orixe evolutivu d'esta proteína puede rastrexase nes célules procariotas, onde esisten equivalentes. D'últimes ye importante nel control de la espresión xénica.
Un bon númberu d'enfermedaes tienen como base alteraciones xenétiques en apanfilos de los xenes que gobiernen la producción de la actina o de les sos proteínes acomuñaes, siendo tamién esencial nel procesu d'infeición de dalgunos microorganismos patóxenos. Les mutaciones nos distintos xenes d'actina presentes n'humanos causen miopatíes, variaciones nel tamañu y la función cardiaca y sordera. Los componentes del citoesqueleto tamién tienen rellación cola patoxenicidá de bacteries intracelulares y virus, especialmente en procesos rellacionaos col fuximientu de la respuesta del sistema inmune.[4]
Historia

L'actina foi reparada esperimentalmente per primer vegada en 1887 por W.D. Halliburton, quien estrayxo una proteína muscular que coagulaba preparaciones de miosina, denominándolo "formientu de la miosina".[5] Sicasí, Halliburton foi incapaz d'efeutuar la carauterización de les sos observaciones, y por ello'l descubrimientu atribuyir a Brúnó F. Straub, entós un mozu bioquímicu que trabayaba nel llaboratoriu d'Albert Szent-Györgyi nel Institutu de química médica de la Universidá de Szeged, n'Hungría.

En 1942, Straub desenvolvió una nueva téunica pa la estraición de proteínes musculares que-y dexaba aisllar cantidaes sustanciales d'actina relativamente pura. Esti métodu ye'l mesmu qu'esencialmente s'utiliza nos llaboratorios anguaño. Szent-Györgyi describiera primeramente una forma más mafosa de miosina, producida por estraiciones lentes en músculu, como "miosina activada" y yá que la proteína de Straub producía l'efeutu activador, denominar actina. La mafa menguaba si añadíase ATP al amiestu de dambes proteínes, conocida como actomiosina. El trabayu de dambos nun pudo ser publicáu nos países occidentales debíu al ambiente bélico de la Segunda Guerra Mundial, saliendo a la lluz en 1945 cuando foi publicáu como suplementu de Acta Physiologica Scandinavica.[7] Straub siguió trabayando na actina hasta 1950, publicando que podía xunise al ATP y que, mientres la polimerización de la proteína pa formar microfilamentos, s'hidrolizaba a ADP + Pi, que permanecía xuníu al filamentu. Straub suxurió qu'esta reaición desempeñaba un papel na contraición muscular, pero esto namái ye ciertu nel casu del músculu llisu y nun foi verificáu esperimentalmente hasta 2001.[8][9]
La secuencia d'aminoácidos foi completada por Elzinga y collaboradores en 1973,[10] y la estructura cristalográfica de la actina G foi determinada en 1990 por Kabsch y collaboradores, anque se trataba d'un cocristal nel que formaba un complexu cola desoxirribonucleasa I,[11] siendo propuestu un modelu'l mesmu añu pa la actina F por Holmes y los sos collaboradores.[12] Esti procedimientu de cocristalización con distintes proteínes foi emplegáu repetidamente mientres los siguientes años, hasta qu'en 2001 llogróse cristalizar la proteína aisllada xunto con ADP. Foi posible gracies al emplegu d'un conxugáu de rodamina que torgaba la polimerización bloquiando l'aminoácidu cys-374.[13] Esi mesmu añu produció'l fallecimientu de Christine Oriol-Audit, la investigadora qu'en 1977 consiguió cristalizar per primer vegada la actina n'ausencia de ABP's. Los cristales resultaron demasiao pequeños pa la teunoloxía de la dómina.[14]
Anque anguaño nun esiste un modelu d'altu resolución de la forma filamentosa, l'equipu de Sawaya realizó en 2008 un aproximamientu más exactu basándose en múltiples cristales de dímeros d'actina que contauten en distintos llugares.[15] Esti modelu foi refináu por el mesmu autor y por Lorenz. Otros enfoques, como l'usu de criomicroscopía electrónica o radiación sincrotrón dexaron apocayá aumentar el nivel de resolución y entender con mayor fondura la naturaleza de les interaiciones y los cambeos conformacionales implicaos na formación del filamentu d'actina.[16][17]
Estructura
L'actina ye una de les proteínes más abondoses ente los eucariotes y atópase presente en tou la citoplasma.[3] Ello ye que nes fibres musculares representa'l 20% en pesu de proteína celular total y, n'otres célules animales, bazcuya ente'l 1 y el 5%. Sicasí, nun esiste un únicu tipu d'actina, sinón que los sos xenes codificantes atópense definíos por una familia multigénica (familia que, en plantes, alluga más de 60 elementos, ente xenes y pseudogenes y, n'humanos, más de 30).[18] Esto significa que la información xenética de cada individuu tien instrucciones pa xenerar variantes de la actina (denominaes isoformes) que van tener funciones llixeramente distintes. D'esta miente, los organismos eucariotes espresen distintos xenes que dan llugar a: la actina α, que s'atopa n'estructures contráctiles; la actina β, nel cantu n'espansión de les célules qu'empleguen la proyeición d'estructures celulares como métodu de movilidá; y la actina γ, nos filamentos de les fibres de estrés.[19] Amás de les semeyances esistentes ente les isoformas d'un organismu, tamién esiste una caltenimientu evolutivu tocantes a estructura y función ente organismos d'inclusive dominios distintos al eucariota: en bacteries conozse l'homólogu MreB, una proteína que ye capaz de polimerizar en microfilamentos;[17] y n'arquies esiste un representante (Ta0583) entá más similar a les actines d'eucariotes.[20]
L'actina presentar na célula en dos formes: como monómeros globulares denominaos actina G y como polímeros filamentosos denominaos actina F (esto ye, filamentos compuestos d'ensame de monómeros d'actina G). L'actina F puede denominase tamién filamentu. A cada hebra d'actina xúnese una molécula d'adenosín trifosfato (ATP) o d'adenosín difosfato (ADP) de la mesma acomuñada a un catión Mg2+. De les distintes combinaciones posibles ente les formes d'actina y el nucleótido trifosfato, na célula predominen la actina G-ATP y la actina F-ADP.[21][22]
Actina G
Tocantes a la so estructura molecular, la actina G tien una apariencia globular al microscopiu electrónicu de barríu; sicasí, por aciu cristalografía de rayos X puede apreciase que ta compuesta de dos lóbulos separaos por una hendidura; la estructura conforma'l plega ATPasa, un centru de catálisis enzimática capaz de xunir el ATP y Mg2+ y hidrolizar el primeru a ADP más fosfatu. Esta plega ye un motivu estructural calteníu que tamién ta presente n'otres proteínes que interaccionan con nucleótidos trifosfato como la hexoquinasa (una enzima del metabolismu enerxéticu) o les proteínes Hsp70 (una familia de proteínes que contribúin a qu'otres proteínes tengan estructures funcionales).[23] L'actina G namái ye funcional cuando tien o bien ADP o bien ATP na so hendidura; sicasí, na célula predomina l'estáu xuníu a ATP cuando la actina atópase llibre.[21]
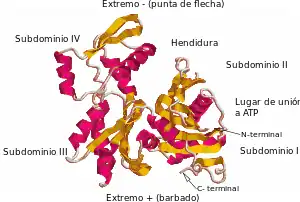
L'actina cristalizada por Kabsch, que ye la más utilizada como modelu n'estudios estructurales, yá que foi la primera en ser purificada, procede del músculu esqueléticu de coneyu. Tien unes dimensiones averaes de 67 x 40 x 37 Å, un masa molecular de 41785 Da y un puntu isoeléctrico envaloráu en 4,8. El so carga neta a pH= 7 ye de -7.[24] [25]
- Estructura primaria
La secuencia d'aminoácidos completa d'esti tipu d'actina foi determinada por Elzinga y collaboradores en 1973, y afinada en trabayos posteriores pol mesmu autor. Contién 374 residuos d'aminoácidos. El so estremu N-terminal ye bien ácidu. Empieza con un aspartato acetiláu nel so grupu amino, ente que'l so C-terminal ye básicu, formáu por una fenilalanina precedida por una cisteína de cierta importancia funcional. Dambos estremos asitiar nuna posición bien próxima dientro del subdominiu I. Tocantes a aminoácidos anómalos, cabo destacar una Nτ-metilhistidina en posición 73.[24]
- Estructura terciaria-dominios
Ta formada por dos dominios conocíos como grande y pequeñu, dixebraos por una hendidura en que'l so centru asitiar el llugar d'unión al ATP-ADP+Pi. Per debaxo d'ésti esiste una escotadura de menor fondura llamada "riegu". Cuando s'atopen en forma nativa, a pesar del so nome, dambos tienen un tamañu equiparable.[10]
Nos estudios topolóxicos, por convención, la proteína empobínase de manera que'l dominiu mayor queda a la izquierda, ente que'l menor asítiase a la derecha. Nesta posición, el dominiu pequeñu estrémase de la mesma nel subdominiu I (posición inferior, residuos 1-32, 70-144 y 338-374) y subdominiu II (posición cimera, residuos 33-69). El dominiu mayor tamién s'estrema n'otros dos, el subdominiu III (inferior, residuos 145-180 y 270-337) y el subdominiu IV (cimeru, residuos 181-269). La zona espuesta de los subdominios I y III denominar estremu "barbáu", ente qu'a la de los subdominios II y IV llámase-y estremu "en punta de flecha". Esta denominación fai referencia al fechu de que por cuenta de la pequeña masa del subdominiu 2, la actina adquier polaridá, que se va aldericar darréu al falar de la dinámica d'ensamblaxe. Dellos autores nomen los subdominios como Ia, Ib, IIa y IIb, respeutivamente.[26]
- Otres estructures destacaes
- La estructura supersecundaria más destacada ye una β-llámina de cinco cadenes que se componen d'un β-meandro y una unidá β-α β dextrógira. Ta presente en dambos dominios. Esto suxure que la proteína surdió por duplicación xénica.[11]
- El llugar d'unión al adenosín nucleótido atopar ente dos estructures en forma de forqueta β pertenecientes a los dominios 1 y 3. Los residuos implicaes son Asp11-Lys18 y Asp154-His161 respeutivamente.
- Xusto debaxo del nucleótido atópase'l llugar d'unión al catión divalente, que in vivo ye con mayor probabilidá'l Mg2+ o'l Ca2+ ente que in vitro ye'l formáu por una estructura quelante na que contribúin la Lys18 y dos osíxenos de los fosfatos α y β del nucleótido. Esti calciu ta coordináu con seis molécules d'agua que s'atopen reteníes polos aminoácidos Asp11, Asp154, y Gln137. Xunto col nucleótido forma un complexu qu'acuta los movimientos d'una rexón llamada gonciu o hinge, asitiada ente los residuos 137 y 144, calteniendo d'esta manera la forma nativa de la proteína, hasta'l puntu de que la so retirada desnaturaliza el monómero d'actina. Esta rexón tamién ye importante, porque determina les conformances "abierta" o "zarrada" de la hendidura de la proteína.[26][13]
- Con cuasi toa probabilidá esisten siquier otros trés centros con menor afinidá (entemedia) y otros de baxa afinidá pa cationes divalentes. Especulóse sobre'l papel d'estos centros na polimerización de la actina actuando na etapa d'activación.[26]
- Nel subdominiu 2 esiste una estructura, llamada bucle-D o D-loop por cuenta de que xúnese a l'ADNasa I, asitiada ente los residuos His40 y Gly48 qu'apaez como un elementu desordenáu na mayoría de los cristales y como una llámina β cuando ta formando complexu cola ADNasa I. Según Domínguez et al., l'eventu clave de la polimerización sería l'espardimientu d'un cambéu conformacional dende'l centru d'unión al nucleótido hasta esti dominiu, que pasaría de ser un bucle a una héliz. Esta teoría paez ser refutada por otros trabayos.[13][27]
Actina F
Una descripción clásica afirma que la actina F tien una estructura filamentosa interpretable como una héliz levógira monocatenaria con xiru de 166º y medría de 27,5 Å o bien como una héliz dextrógira bicatenaria con mediu pasu de rosca de 350-380 Å, tando cada actina arrodiada d'otros cuatro.[28] La simetría del polímeru d'actina, que ye d'unes 2,17 subunidades por vuelta de héliz ye incompatible cola formación de cristales, que namái ye posible cuando éstes son esautamente 2, 3, 4 ó 6 subunidades por vuelta. Por tanto, débense efeutuar modelos interpretando datos procedentes de téuniques que salven estos inconvenientes, como la microscopía electrónica, la criomicroscopía electrónica, cristales de dímeros en distintes posiciones o difraición de rayos X.[17] Ye necesariu precisar que falar d'una "estructura" nun ye correutu pa daqué tan dinámicu como un filamentu d'actina. En realidá tendría de falase de distintos estaos estructurales, ente los cualos el datu más constante ye la medría de 27,5 Å, ente que la rotación de les subunidades amuesa una considerable variabilidá, siendo normal reparar desplazamientos d'hasta'l 10% de la so posición ideal. Delles proteínes, como la cofilina, paecen amontar l'ángulu de xiru, pero nuevamente puede interpretase que, en llugar d'ello, estabilicen dellos "estaos estructurales" normales. Estos podríen ser importantes nel procesu de polimerización.[29]
En cuanto al radiu de xiru o grosez del filamentu, les midíes son más revesoses: mientres los primeros modelos asignáben-y una llargor de 25 Å, datos actuales de difraición de rayos X sofitaos por criomicroscopía electrónica coinciden nunos 23,7 Å. Estos mesmos estudios determinaron con bastante precisión los puntos de contautu ente monómeros. Unos establécense con unidaes de la mesma cadena, ente l'estremu "barbáu" d'un monómero y l'estremu "en punta de flecha" del siguiente, ente que los monómeros de cadenes axacentes faen contautu lateralmente por aciu proyeiciones del subdominiu 4, siendo les más importantes la formada pol C-terminal y un enllaz hidrofóbicu formáu por trés cuerpos nos qu'intervienen los residuos 39-42, 201-203 y 286. Pa formar parte d'un filamentu, según esti modelu, los monómeros taríen nuna configuración llamada "plana", na que los subdominios xiren ente sigo, y que tamién paez atopase nel homólogu bacterianu de la actina MreB.[17]
Puesto que toles subunidades d'un filamentu apunten escontra'l mesmu estremu, dizse que'l polímeru presenta polaridá na so estructura. Esti fechu da llugar a una convención: nomar al estremu que tien una subunidad d'actina esponiendo'l llugar pol que xune ATP al mediu como «estremu (-)» ente que nel opuestu, nel cual la hendidura ta dirixida a otru monómero axacente, ye'l «estremu (+)».[19] La denominación «en punta de flecha» y «barbáu» de los estremos de los microfilamentos deber al so aspeutu al microscopiu electrónicu de tresmisión cuando se procesen por aciu una téunica denominada «decoración». Esti métodu consiste na adición d'elementos S1 de la miosina en texíos fitos con acedu tánico; esta miosina xune de forma polar a los monómeros d'actina, lo que da llugar a una configuración asemeyada a fleches con plumes a lo llargo de tol so fuste, onde'l fuste correspondería a la actina y les plumes a la miosina. D'esta miente, l'estremu del filamentu que queda ensin miosina sobresaliendo interprétase como la punta de la flecha, mientres l'opuestu denominar barbáu.[30]
Nel músculu, el filamentu helicoidal de la actina F contién tamién una molécula de tropomiosina, una proteína d'un llargor de 40 nanómetros que s'endolca alredor de la héliz d'actina F. Mientres l'estáu de reposu celular, la tropomiosina anubre los sitios activos de la actina de cuenta que nun se llogra la interaición actina-miosina que produz la contraición muscular. Xuníes a lo llargo de la hebra de tropomiosina hai otres molécules proteiques, les troponines, complexos de tres polímeru: troponina I, troponina T y troponina C.[31]
Plegamientu
L'actina puede adquirir bonalmente una gran parte de la so estructura terciaria.[33] Sicasí, amuesa un comportamientu bien especial y cuasi únicu na forma en qu'adquier la so forma dafechu funcional a partir de la so forma nativa recién sintetizada. La razón d'una ruta tan especial podría ser la necesidá d'evitar la presencia de monómeros d'actina mal plegaos, que seríen tóxicos yá que podríen actuar de terminadores desaparentes de la polimerización. Sía que non, ye clave pa la estabilidá del citoesqueleto, y non yá esto, sinón que podría ser un procesu esencial pa la coordinación del ciclu celular.[34][35]
Pa ello emplega obligadamente un tipu de chaperonina (proteína qu'ayuda a otres a plegase) citosólica del grupu II, la CCT, formada por un doble aniellu d'ocho subunidades distintos (heteroctamérico) que s'estrema de les demás chaperonas moleculares, y n'especial de la so homóloga n'arquies GroEL en que nun precisa d'una co-chaperona qu'actúe de tapadoria sobre'l cuévanu central catalítica. Acepta sustratos xuniéndose a ellos por aciu dominios específicos, polo qu'en principiu se pensó que yera esclusiva d'actina y tubulina, anque anguaño viose por inmunoprecipitación que siquier interactúa con un gran númberu de polipéptidos, posiblemente como sustratos. Actúa por aciu cambeos conformacionales dependientes de ATP, precisando n'ocasiones de delles rondes de lliberación y catálisis pa completar el so trabayu.[36]
Pal so correutu plegamientu, la actina y la tubulina tamién precisen específicamente del concursu d'otra proteína, la prefoldina, un complexu heterohexamérico (formáu por seis subunidades distintes), y tan específicamente qu'inclusive han coevolucionado. Nel casu de la actina, xúnese darréu a ella mientres entá se ta traduciendo, aproximao cuando tien un llargor de 145 aminoácidos, que son los correspondientes al dominiu N-terminal.[37]
Empléguense subunidades de reconocencia distintes pa la actina y la tubulina, anque asolapaes. Probablemente nel casu de la actina tratar de les subunidades PFD3 y PFD4 que se xunen a la actina en dos llugares, el I, ente los residuos 60–79 y el II, ente los residuos 170–198. L'actina reconozse, carga y entrega a la CCT en conformanza abierta pela parte interna del estremu de los "tentáculos" de la prefoldina (ver imaxe y nota al pie).[n. 1] El contautu nel momentu de la entrega ye tan curtia que nun se llega a formar un complexu ternariu, lliberar la prefoldina de momentu.[32]

Darréu, la chaperonina citosólica (CCT) efectúa'l plegamientu de la actina de forma secuencial, y formando uniones coles subunidades, en llugar d'a cencielles zarrala nel so cuévanu.[n. 2] Pa ello tien zones específiques de reconocencia nel so dominiu apical. La primer etapa del plegamientu consistiría na reconocencia de los residuos 245-249. Darréu, otros determinantes estableceríen contautu.[38] Tanto la actina como la tubulina xunir a la CCT en conformances abiertes n'ausencia de ATP. Nel casu de la actina, en cada cambéu conformacional xunir a dos subunidades, a diferencia de la tubulina, que lo fai a cuatro. L'actina tien secuencies d'unión específiques, interactuando coles subunidades CCTδ y β o bien con CCTδ y CCTε. Tres la unión de AMP-PNP a la CCT, los substratos van moviéndose pol cuévanu de la chaperonina. Paez ser tamién que nel casu de la actina precísase la proteína CAP como un posible cofactor nos estadios finales del plegamientu de la actina.[35]
Entá nun se conoz con exactitú la regulación d'esti procesu, pero sábese que la proteína PhLP3 (proteína asemeyao a la fosducina) regula la so actividá tornándola, por aciu la formación d'un complexu ternariu.[36]
Mecanismu catalíticu de la ATPasa
L'actina ye una ATPasa, esto ye, una enzima qu'hidroliza ATP. Esti conxuntu d'enzimes carauterizar por actuar con estrema lentitú. Sábese qu'esta ATPasa se "activa", o lo que ye lo mesmo, la so velocidá aumenta unes 40 000 vegaes cuando la actina forma parte d'un filamentu.[29] Un valor de referencia pa esta tasa d'hidrólisis so ciertes condiciones ideales sería de 0,3 s-1. Darréu, el Pi permanecería llargu tiempu xuníu a la actina xunto al ADP, lliberar cerca del estremu del filamentu.[39]
A día de güei nun se conocen los detalles moleculares concretos del mecanismu catalíticu. Anque esiste muncho discutiniu al respeutu, paez claro que pa la hidrólisis de ATP precísase una conformanza "zarrada", y créese qu'avera los residuos implicaes a la distancia fayadiza.[29] Unu de los residuos clave sería Glu137, asitiáu nel subdominiu 1. La so función sería fondiar la molécula d'agua que produz un ataque nucleofílico al enllaz del fosfatu γ del ATP, mientres el nucleótido xúnese fuertemente a los subdominios 3 y 4. La lentitú del procesu catalíticu deber a la gran distancia y posición sesgada d'esta molécula d'agua con al respective de la so reactante. Con muncha probabilidá, el cambéu conformacional que se produz por rotación de dominios ente les formes G y F de la actina avera la Glu137, dexando la so hidrólisis. Según esti modelu, la polimerización y la función ATPasa taríen desacopladas nun primer momentu.[17]
Dinámica d'ensamblaxe

L'actina F combina les cualidaes de ser resistencia de materiales resistente y dinámica. A diferencia d'otros polímeros, como l'ADN, que caltienen xuníos los sos elementos constitutivos por aciu enllaces covalentes, nos filamentos d'actina los monómeros se ensamblan por enllaces más débiles de tipu non covalente. Esto, qu'en principiu debilita la estructura, yá que podría rompese por baturiciu térmicu, solucionar por aciu los enllaces llaterales colos monómeros vecinos. Coles mesmes, los enllaces débiles caltienen la ventaya de que los estremos del filamentu pueden lliberar o incorporar fácilmente monómeros, de manera que pueden remocicase rápido y camudar la estructura celular de la que son responsables en respuesta a estímulos ambientales. Esto últimu y el mecanismu bioquímicu pol que s'efectúa ye lo que se conoz como "dinámica d'ensamblaxe".[4]
- Estudios in vitro
Los estudios de la dinámica de adición y perda de subunidades de los microfilamentos realizáronse in vitro (esto ye, nel llaboratoriu, fora de sistemes celulares) por cuenta de que'l polímeru d'actina resultante da llugar a la mesma actina F producida in vivo, onde esti procesu ta controláu por ensame de proteínes pa responder a les necesidaes celulares, de cuenta que sería bien difícil reparar les sos condiciones básiques.[40] In vitro, esti fechu producir de forma secuencial: primero, dase una fase d'activación», na que la unión ya intercambiu de cationes divalentes en llugares específicos de la actina G, xuníu a ATP, producen un cambéu conformacional, conocíu dacuando como Actina G* o monómero d'actina F, yá que ye más asemeyada a les unidaes que s'asitien nel filamentu.[26] Esto preparar pa la siguiente «fase de nucleación», na cual la actina G da llugar a pequeños fragmentos inestables d'actina F capaz de polimerizar. Primeramente fórmense dímeros y trímeros de manera inestable. Cuando'l númberu d'estos ye lo suficientemente grande, tien llugar la fase de elongación», na que'l filamentu fórmase y crez rápido por aciu la adición reversible de nuevos monómeros a dambos estremos.[41] Finalmente, nel equilibriu estacionariu», los monómeros d'actina G intercambiar nos estremos del filamentu ensin que varie'l llargor total del polímeru.[3] Nesta última fase defínese la concentración crítica Cc» como la rellación ente les constantes d'ensamblaxe y desensamblaje (trátase, pos, d'una constante de disociación), y representa la concentración d'actina G na cual la dinámica de adición y eliminación de monómeros nun produz un cambéu nel llargor del filamentu. Nes condiciones avezaes in vitro, Cc ye de 0,1 μM,[42] lo que significa qu'a valores mayores dase una polimerización y a valores menores, una despolimerización.[43]
- Papel de la hidrólisis de ATP
Un asuntu importante que s'introdució nel apartáu anterior ye'l fechu de que, anque la actina hidroliza ATP, tou paez indicar qu'esto nun intervién nel ensamblaxe, yá que, per una parte, la hidrólisis producir en gran midida nel interior del filamentu, y por otra, el ADP tamién puede polimerizar. Esto plantega la cuestión d'entender cuál ye'l procesu termodinámicamente desfavorable que rique un gastu d'enerxía tan ingente. El llamáu "ciclu de la actina", qu'amiesta la hidrólisis a la polimerización, consiste na adición de monómeros d'Actina G-ATP preferentemente nel estremu barbáu, creando un fluxu de monómeros escontra l'estremu en punta de flecha no que se conoz como "trenzamiento" (threadmilling, n'inglés), onde los monómeros taríen en forma d'Actina F-ADP y seríen lliberaos, intercambiando darréu esti ADP por ATP y cerrando d'esa manera'l ciclu.
Poco dempués de la adición, produzse la hidrólisis del ATP de forma relativamente rápida. Esisten dos hipótesis sobre cómo se produz, la estocástica, na que la hidrólisis producir al azar influyida en ciertu mou poles molécules vecines, y la vectorial, na que namái se produciría na llende con otres molécules que yá han hidrolizado la so ATP. Sía que non, nun se lliberar el Pi resultante, sinón que permanez un tiempu xuníu non covalentemente a la actina ADP, colo cual esistiríen tres especies d'actina nun filamentu: ATP-Actina, ADP+Pi-Actina y ADP-Actina.[39] El conteníu d'un filamentu en caúna d'estes especies depende del so llargor y estáu: al empiezu de la elongación, el filamentu tien una composición aproximao equivalente de monómeros con ATP y ADP+Pi y una pequeña cantidá xunto al estremu (-) d'Actina ADP. A midida qu'algámase l'estáu estacionariu, la situación inviértese, tando la mayor parte del filamentu con ADP y l'estremu (+) práuticamente namái con ADP+Pi, col ATP amenorgáu al estremu.[44]
Si comparamos los filamentos d'actina-ADP puros con aquellos qu'incorporen ATP, nos primeres les constantes crítiques son similares en dambos estremos, ente que nos otros dos nucleótidos la Cc ye distintu: Nel estremu (+) ye Cc+=0.1 μM, ente que nel estremu (-) ye Cc-=0.8 μM, colo cual danse les siguientes situaciones:[19]
- Pa concentraciones d'actina G-ATP menores a Cc+ nun se produz la elongación del filamentu.
- Pa concentraciones d'actina G-ATP menores que Cc- pero mayores que Cc+ la elongación dar nel estremu (+).
- Pa concentraciones d'actina G-ATP mayores que Cc- el filamentu crez en dambos estremos.
Por tanto, puede deducise que la enerxía de la hidrólisis utilizar pa crear un verdaderu "estáu estacionariu", esto ye, d'un fluxu en llugar d'un simple equilibriu, lo cual dota de dinamismu, polaridá y fuercia de traición al filamentu, lo que xustifica'l gastu pola ganancia de funciones biolóxiques esenciales.[39] Amás, la configuración de los distintos tipos de monómeros ye detectada poles proteínes d'unión a la actina que controlen esti dinamismu, como se va ver na próxima seición.
Paez esistir una esceición na forma d'acoplamientu típica de los microfilamentos por aciu trenzado o threadmilling nos esterocilios. Nesti casu, el control del tamañu de la estructura sería totalmente apical y de dalguna manera controlada pola espresión xénica, esto ye, pola cantidá total de monómero de proteína sintetizada nun momentu dau.[45]
Proteínes acomuñaes

In vivo, el citoesqueleto d'actina nun ta compuestu puramente d'actina, sinón que pa la so xeneración, permanencia y función rique d'otres proteínes; éstes denominar proteínes d'unión a la actina (ABP, actin binding proteins) ya intervienen nel so polimerización y despolimerización, estabilidá, la so organización en fexes o redes, la so fragmentación y destrucción.[3] La diversidá d'estes proteínes ye tal, que se considera que la actina ye la proteína que participa nel mayor númberu d'interaiciones proteína-proteína de cuantes conócense.[47] Por casu, esisten elementos que secuestren la actina G, torgando la so incorporación a los microfilamentos. Otramiente, esisten proteínes qu'aguiyen la so polimerización o que doten de complexidá a les redes en resume.[19]
- La timosina β4 ye una proteína de 5 kDa capaz de xunise a la actina G-ATP nuna estequiometría 1:1; esto quier dicir qu'una unidá de timosina β4 xunir a otra d'actina G, nesta proporción. El so papel ye torgar la incorporación de los monómeros al polímeru en crecedera.[48]
- La profilina, una proteína citosólica de 15 kDa que tamién se xune en estequiometría 1:1 a los monómeros d'actina G-ATP, pero la so función ye distinta: facilita l'intercambiu de nucleótidos ATP por ADP. Amás, ta implicada n'otres funciones celulares, como la unión de repeticiones de Pro n'otres proteínes o de lípidos qu'actúen como segundos mensaxeros.[49][50]

Otres proteínes d'unión a actina regulen el llargor de los microfilamentos realizando cortes nellos, lo que da llugar a nuevos estremos activos pa la polimerización. Esto ye, si un filamentu, que tien dos estremos a los que pueden xunise o disociarse monómeros, ye cortáu dos vegaes, resulten trés nuevos microfilamentos con seis estremos; la nueva situación favorez la dinámica d'ensamblaxe y desensamblaje. Ente estes proteínes destaquen la gelsolina y la cofilina. Cabo resaltar que primero realicen la corte por aciu cambeos na conformanza del monómero d'actina al que se xunen nel polímeru, quedando dempués anubriendo'l nuevu estremu (+) xeneráu, lo que torga l'agregáu o l'intercambiu de nueves subunidades d'actina G y, yá que los estremos (-) queden ensin anubrir, favorecen la despolimerización de los filamentos.[52]
Otru tipu de proteínes d'unión a actina anubren los estremos de la actina F con cuenta d'estabilizalos, ensin capacidá de rompelos. Exemplos d'estes proteínes son CapZ (que xune los estremos (+) según los niveles de Ca2+/calmodulina de la célula, niveles que dependen de señales esternes ya internes de la célula y qu'intervienen na regulación de les sos funciones biolóxiques)[53] o la tropomodulina (que xune los estremos (-)). La tropomodulina ye esencial como estabilizador de la actina F presente nes miofibrilles de los sarcómeros del músculu, estructures caracterizaes pol so gran estabilidá.[54]

El complexu Arp2/3 atópase llargamente espublizáu en tolos organismos eucariotes.[56] Ta compuestu por siete subunidades, dalgunes de les cualos tienen una topoloxía claramente rellacionada cola so función biolóxica: dos de los sos subunidades, denominaes «ARP2» y «ARP3», tienen una estructura bien asemeyada a los mesmos monómeros d'actina. Dicha homoloxía dexa a dambes unidaes portase como axentes nucleantes de la polimerización de los monómeros d'actina G a actina F. Amás, esti complexu ye necesariu pa establecer estructures dendríticas y en anastomosis (esto ye, encruciaes o en rede), por tanto más complexes, d'actina F.[57]
Inhibidores químicos
Esisten delles toxines qu'interfieren cola dinámica de les actines, tantu despolimerizándolas (latrunculina y citocalasina D) como estabilizándoles (faloidina):
- La latrunculina, una toxina producida por poríferos, xunir a la actina G torgando la so unión a los microfilamentos.[58]
- La citocalasina D, un alcaloide producíu por fungos, xunir al estremu (+) de la actina F torgando la adición de nuevos monómeros.[59] Describiéronse efeutos de la citocalasina D, mediaos pola disrupción de la dinámica d'actines, na actividá de p53 (n'animales)[60] o en respuestes gravitrópicas (en plantes).[61]
- La faloidina, una toxina aisllada del fungu Amanita phalloides, xunir a la interfaz esistente ente los monómeros d'actina axacentes del polímeru d'actina F, lo qu'evita la despolimerización d'aquél.[59]
Funciones y llocalización
L'actina como proteína atópase tantu nel citoplasma como nel nucleu celular.[62] Felicidad llocalización ta regulada peles víes de transducción de señales qu'integren los estímulos que la célula recibe y que dexa la reestructuración de les redes d'actina en respuesta a aquellos. En Dictyostelium, refirióse la intervención de la ruta de fosfoinosítidos mediada pola fosfolipasa D.[63] Los filamentos d'actina son especialmente abondosos y estables nes fibres musculares. Dientro del sarcómero (la unidá morfolóxico y fisiolóxico de les fibres musculares) la actina disponer nes bandes I y A; nesta postrera, preséntase conxuntamente cola miosina.[64]
Citoesqueleto
Los microfilamentos intervienen nel movimientu de toles célules móviles, inclusive les non musculares, pos se describió que los fármacos que desorganizan la actina F (como les citocalasines) afecten a l'actividá de diches célules. Como proteína, la actina supón el 2% del total de proteínes en hepatocitos, el 10% en fibroblastos, el 15% n'amebes y hasta'l 50-80% en plaquetes activaes.[65] Esisten distintos grupos d'actina, con estructura y función llixeramente distintes. D'esta miente, la actina α ye esclusiva de fibres musculares, y el presente n'otres célules suel ser del tipu β y γ. Amás, la actina de tipos distintos a soler tener una alta tasa de recambiu que provoca que la mayor parte d'ella nun forme parte d'estructures permanentes. Asina, los microfilamentos nes célules non musculares apaecen de dos formes:[66]
- Redes de microfilamentos. So la membrana plasmática ye común en célules animales l'apaición d'una corteza celular poblada por ensame de microfilamentos qu'esclúi la presencia d'orgánulos. Esta rede ta en rellación con abondoses receptores celulares que transducen señales del esterior de la célula.
- Fexes de microfilamentos. Estos microfilamentos, dispuestos en redes, son de mayor llargor y, n'asociación con proteínes contráctiles como la miosina non muscular, intervienen nel desplazamientu de sustancies a nivel intracelular.
Lleldos
En lleldos, el citoesqueleto d'actina ye clave mientres los procesos d'endocitosis, citocinesis, determinación de la polaridá celular y mientres la morfogénesis. Estos fechos, amás de depender de la actina, impliquen a 20 o 30 proteínes acomuñaes, altamente calteníes evolutivamente, según ensame de molécules de señalización; estos elementos dexen, en combinación, un ensamblaxe espacial y temporalmente modulado que define la bioloxía celular en respuesta a estímulos internos y esternos.[67]
Los lleldos tienen trés grandes tipos d'elemento producto de l'asociación de la actina: parches, cables y aniellos que, magar detectase mientres llargos periodos de tiempu, vense sometíos a un equilibriu dinámicu por cuenta de la continua polimerización y despolimerización. Como proteínes accesories, tienen una cofilina/ADF de 16 kDa (codificada por un únicu xen, denomináu COF1); Aip1, un cofactor de la cofilina que favorez el desensamblado de los microfilamentos; Srv2/CAP, un regulador de la dinámica rellacionáu con proteínes adenilil ciclasas; una profilina d'aproximao 14 kDa que s'acomuñar a los monómeros d'actina; y twinfilina, una proteína de 40 kDa implicada na organización de les estructures tipu parche.[67]
Plantes
Los estudios de xenómica de plantes revelaron la esistencia de isovariantes proteiques dientro de la familia de xenes de la actina; dientro de Arabidopsis thaliana, una dicotiledónea emplegada como organismu modelo, esisten siquier diez tipos d'actines, nueve de α tubulinas, seis de β tubulinas, seis de profilinas y docenes de miosinas. Tal diversidá esplícase acordies cola necesidá evolutiva de tener variantes llixeramente distintes na so pauta d'espresión temporal y espacial; sicasí, la mayoría d'elles esprésense conxuntamente nos texíos analizaos. El treme de redes d'actina distribuyir por tou la citoplasma de les célules cultivaes in vitro, con un refuerzu en redol al nucleu que se coneuta, por aciu radios, a la corteza celular; dichu treme ye altamente dinámicu, con un polimerizado y despolimerizado continuu.[68]

Magar les célules vexetales tienen xeneralmente una paré que define la so morfoloxía y torga el so movimientu, los sos microfilamentos xeneren les fuercies necesaries pa delles actividaes celulares, por casu, les corrientes citoplasmátiques xeneraes polos microfilamentos y les miosines. Amás, la actina intervién nel movimientu de orgánulos y morfogénesis celular, procesos qu'inclúin la división celular, la elongación y la diferenciación.[70]
Tocantes a les proteínes acomuñaes al citoesqueleto d'actina presentes en plantes cabo mentar:[70] la villina, una proteína perteneciente a la familia de la gelsolina/severina, capaz de cortar microfilamentos y xunir monómeros d'actina en presencia del catión calciu; la fimbrina, un elementu capaz de reconocer y xunir monómeros d'actina y qu'intervién na formación de tremes (por aciu una regulación distinta a la mesma de célules animales y lleldos);[71] les formines, proteínes capaces d'actuar como axente nucleante de la polimerización a actina F; la miosina, típicu motor molecular propiu d'eucariotes que, en Arabidopsis thaliana, ta codificado por 17 xenes clasificaos en dos clases distintes; CHUP1, capaz de xunir actina ya implicáu na distribución espacial de los cloroplastos na célula; KAM1/MUR3, una proteína que define la morfoloxía del complexu de Golgi según la composición en xiloglucanos de la paré celular; NtWLIM1, proteína que faculta l'apaición d'estructures aovilladas d'actina; y ERD10, que participa na asociación ente orgánulos delimitados por membranes y los microfilamentos y que paez desempeñar un papel especialmente relevante en presencia d'estrés.
Contraición muscular

Nel músculu, el filamentu helicoidal de la actina F contién tamién una molécula de tropomiosina, una proteína d'un llargor de 40 nanómetros que s'endolca alredor de la héliz d'actina F. Mientres l'estáu de reposu celular, la tropomiosina anubre los sitios activos de la actina de cuenta que nun se llogra la interaición actina-miosina (esta interaición da llugar a un deslizamiento ente dambos que, por coordinación de munches copies d'estos elementos dispuestos nos músculos, produz la so contraición). Xuníes a lo llargo de la hebra de tropomiosina hai otres molécules proteiques, les troponines, complexos de tres polímeru: troponina I, troponina T y troponina C.[31] La función moduladora de la tropomiosina depende de la interaición cola troponina en presencia d'iones de Ca2+.[72]
L'actina, xunto cola miosina, intervién na contraición y relaxación de los músculos, constituyendo los dos alredor del 90% de les proteínes musculares.[73] El procesu global disparar por aciu una señal esterna, típicamente por aciu un potencial d'aición excitador del músculu qu'alluga les célules especializaes riques en filamentos d'actina y miosina nel so interior. El ciclu de contraición-relaxación respuende a los siguientes pasos:[74]
- Despolarización del sarcolema y tresmisión del potencial d'aición al traviés de los túbulos T.
- Apertura de canales de Ca2+ del retículo sarcoplásmico.
- Aumentu de la concentración citosólica de Ca2+ ya interaición d'estos cationes cola troponina causando un cambéu nel so conformanza, lo qu'alteria de la mesma la estructura de la tropomiosina, qu'anubre'l sitiu activu de la actina, dexando l'establecimientu de los enllaces cruzaos miosina-actina (esta postrera presente como filamentos delgaos).[31]
- Movimientu de les cabeces de miosina sobre los filamentos delgaos, tantu de forma independiente como dependiente de ATP. Esti últimu mecanismu, mediáu pola actividá ATPasa de les cabeces de miosina, provoca'l movimientu de los filamentos d'actina escontra'l discu Z.
- Captura del Ca2+ per parte del retículo sarcoplásmico, que provoca un nuevu cambéu conformacional na tropomiosina que inhibe la interaición actina-miosina.[73]
Otros procesos biolóxicos
L'estudiu clásicu de la función de la actina la circunscribe al caltenimientu del citoesqueleto y, por ello, a la organización y movimientu de los orgánulos y determinación de la forma celular.[66] Sicasí, el papel de la actina ye abondo más ampliu na fisioloxía celular eucariota; entá más, esisten elementos asemeyaos en procariotes.
- Citocinesis. Nes célules animales y de lleldos, la división celular suel traer la separación de la célula madre en dos célules fíes por aciu la constricción de la so zona ecuatorial. Nesti procesu intervién un aniellu contráctil d'actina, miosina y α actinina.[75] Nel lleldu de fisión Schizosaccharomyces pombe, la actina se ensambla viviegamente nel aniellu contráctil cola participación d'Arp3, la formina Cdc12, profilina y WASp, magar intervienen tamién microfilamentos preformados. Una vegada constituyíu l'aniellu, la estructura caltener nun continuu ensamblado/desensamblado que, cola ayuda del complexu Arp2/3 y de les forminas, apuerta nun procesu central de la citocinesis.[76] El conxuntu d'aniellu contráctil, microtúbulos del fusu acromáticu y el material trupo periféricu denominar «cuerpu de Fleming» o «cuerpu entemediu».[66]
- Apoptosis. Mientres la muerte celular programada, la familia de proteasas denominaes ICE/ced-3 (de la familia de les proteasas conversoras de interleuquina-1β) degraden in vivo la actina en dos fragmentos de 15 kDa y 31 kDa, lo que supón unu de los mecanismos de destrucción de la viabilidá celular en que se basa la apoptosis.[77] Tamién se citó esta destrucción por aciu la proteasa calpaína;[78] tanto ye asina, que l'emplegu de inhibidores de la calpaína mengua la proteólisis de la actina y, inclusive, la degradación del ADN (otru de los elementos carauterísticos de la apoptosis).[79] Per otru llau, la inducción del procesu de apoptosis por aciu un estrés pasa pola reorganización del citoesqueleto d'actina (lo qu'implica tamién el so polimerización), dando llugar a les estructures denominaes fibres de estrés; esti fechu ta señalizáu por aciu la vía de les MAP kinasas.[80]

- Adhesión celular y desarrollu. L'adhesión ente célules ye un calter de los organismos pluricelulares que sofita la capacidá d'especialización tisular y, por ello, l'aumentu de la complexidá d'aquellos. Les uniones celulares de los epitelios empleguen el citoesqueleto d'actina, dientro de cada célula, y les cadherines, como elementos estracelulares, con una conexón ente dambes mediada por catenines.[81] La interrupción de la dinámica d'actines repercute nel desenvuelvo de los organismos; ello ye que la actina ye un elementu tan crucial que, xeneralmente, disponer de sistemes de xenes redundantes. Por casu, los exemplares de Dictyostelium a los que se-yos quitara del xen de la α actinina o del factor gelificante nun amosaben un fenotipu anómalu posiblemente por cuenta de que una de les proteínes podía realizar la función de la otra; sicasí, nos dobles mutantes, carentes de dambes, el desarrollu viose alteriáu.[82]
- Modulación de la espresión xénica. L'estáu de polimerización d'actina inflúi na pauta d'espresión xénica. Nel añu 1997, en trabayos emplegando célules de Schwann detectóse que la despolimerización mediada por citocalasina D provocaba una pauta d'espresión peculiar de los xenes implicaos na mielinización d'esti tipu de célula nerviosa.[83] Tocantes a los organismos unicelulares, en dalguna de les sos fases vitales demostróse que la actina F tamién modifica'l transcriptoma nel fungu Candida albicans.[84] Amás, proteínes asemeyaes a la actina desempeñen un papel regulador mientres la espermatogénesis nel mure[85] y, en lleldos, propúnxose un papel de proteínes asemeyaes a actina na modulación epixenética.[86] Ello ye que la actina ye capaz, xunto con un tipu de miosina nuclear d'interactuar con ARN polimerasas y otres enzimes de la maquinaria transcripcional, d'actuar como iniciador de la trescripción.[62]
- Dinámica d'estereocilios. Dellos tipos de célules desenvuelven na so superficie unes fines evaginaciones filiformes con función mecanosensorial denominaes estereocilios. Por casu, estos orgánulos son los implicaos nel sentíu del oyíu nel órganu de Corti. Como carauterística principal, estes estructures tienen un llargor que puede modificar.[87] Tocantes a la so arquiteutura molecular, los estereocilios tienen un nucleu paracristalino d'actina n'equilibriu dinámicu colos monómeros presentes nel citosol axacente. A lo llargo d'esti nucleu dispónense miosinas de los tipos VI y VIIa, ente que la miosina XVa tar nos sos estremos y en cantidaes proporcionales al llargor del estereocilio.[88]
Patoloxía molecular
Na mayoría de los mamíferos esisten seis xenes distintos d'actina. Dos d'ellos tán rellacionaos col citoesqueleto (ACTB y ACTG1) ente que les cuatro restante tar col músculu esqueléticu (ACTA1), músculu llisu (ACTA2), músculu llisu entérico (ACTG2) y col músculu cardiacu (ACTC1). Les mutaciones qu'afecten a estos xenes yeren desconocíes hasta 1998, y viose que producen miopatíes, variaciones nel tamañu y la función cardiaca y sordera. Asina mesmu, la actina del citoesqueleto ta implicada nel mecanismu de patoxenicidá de múltiples axentes infeiciosos, incluyendo'l VIH. La inmensa mayoría de les mutaciones qu'afecten a la actina son de tipu puntual y tienen un efeutu dominante, salvu siquier seis mutaciones de miopatía nemalínica. Ello ye por cuenta de qu'en munchos casos la variedá mutante del monómero d'actina actúa faciendo de "capping", esto ye, como terminador de la elongación de la actina F.[26]
Rellacionada con ACTA1
ACTA1 ye'l xen que codifica la isoforma α de la actina humana presente principalmente nel músculu esqueléticu, anque tamién s'espresa nel músculu cardiacu y na glándula tiroides.[89] El so secuencia consta de siete exones, que producen cinco transcritos conocíos.[90] A fecha de 2006, el ENMC (European Neuromuscular Centre) publicara 116 mutaciones rellacionaes con patoloxíes, conocíes como actinopatías. La mayor parte d'elles consisten en sustituciones puntuales d'aminoácidos, qu'en munchos casos pueden ser acomuñaes col fenotipu que determina la severidá y el cursu de l'afección.[26][90]
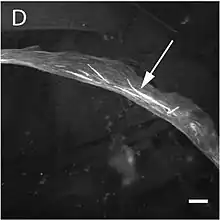
Manifiéstense alteriando la estructura y la función del músculu esqueléticu produciendo trés formes de miopatía: miopatía nemalínica tipu 3, miopatía conxénita con escesu de microfilamentos (CM) y miopatía conxénita con desproporción de tipos de fibra (CFTDM). Tamién se detectó mutaciones que producen miopatía con cores (zones desprovistes d'actividá oxidativa).[92] Anque los sos fenotipos son similares, amás de la miopatía nemalínica típica y la de cayaos intranucleares, dellos especialistes estremen un tipu de miopatía, llamada actínica de la miopatía nemalínica. Na primera atrópense amestaos d'actina en llugar de los típicos cayaos. Ye importante señalar qu'un paciente puede amosar más d'unu d'estos fenotipos na biopsia.[93] Los síntomes más habituales consisten nuna morfoloxía facial típica (facies miopática), debilidá muscular y retrasu nel desarrollu motor y dificultaes respiratories. El cursu, la gravedá y la edá d'apaición son bien variables, y atópense formes de miopatía asolapaes. Na miopatía nemalínica apaecen unes estructures non patognomónicas en diverses llocalizaciones de les fibres musculares tipu 1 conocíes como "cayaos nemalínicos", con una composición similar a los discos z del sarcómero.[94]
La patoxénesis ye bien variada. Munches mutaciones dar na zona de hendidura de la actina, próximes al llugar d'unión pa nucleótidos, ente qu'otres danse en dominiu 2, o bien nes zones d'interaición coles proteínes acomuñaes, lo qu'esplica la gran variedá d'agregaos que se formen nestos casos, como cuerpos nemalínicos, intranucleares o cuerpos zebra.[26] Na miopatía nemalínica prodúcense cambeos nel plegamiento y nes propiedaes d'agregamientu de la actina, y tamién na espresión d'otres proteínes acomuñaes. En delles variantes nes que s'atopen cuerpos intranucleares, el cambéu nel plegamientu despinta la señal d'esportación nuclear, de cuenta que l'agregamientu de la forma mutante d'actina producir nel nucleu celular.[95] Sicasí, paez que nes mutaciones de ACTA1 que dan llugar a CFTDM ta más afeutada la función sarcomérica que la estructura en sí.[96] Trabayos recién traten d'esclariar l'aparente paradoxa de que nun esista una correllación clara ente la bayura de cayaos y la debilidá muscular. Paez ser que delles mutaciones particulares son capaces d'inducir una mayor tasa d'apoptosis nes fibres musculares tipu II.[34]
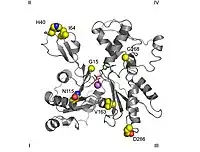
De músculu llisu
Esisten dos isoformas que codifican actines del músculu llisu:
ACTG2 codifica la isoforma más llarga d'actina, con nueve exones, unu d'ellos, l'asitiáu nel estremu 5', que non se traduz.[97] Trátase d'una γ actina que s'espresa nel músculu llisu entérico. Nun s'atoparon mutaciones que se correspuendan a patoloxíes nesti xen, anque se vio por aciu microarrays que ye la proteína que, con diferencia, más aumenta la so espresión nos casos de resistencia a la quimioterapia con cisplatino.[98]
ACTA2 codifica una α actina alcontrada nel músculu llisu, y tamién nel músculu llisu vascular. Viose qu'una mutación, la MYH11, podría ser responsable de siquier un 14% de los casos d'aneurismas d'aorta torácica hereditaria, concretamente'l tipu 6, yá que la variante mutada produz un mal ensamblaxe de los filamentos y un amenorgamientu de la capacidá de contraición del músculu llisu vascular. Reparar nestos individuos dexeneración aórtica medial, con árees de desorganización y hiperplasia, y estenosis de los vasa vasorum de l'aorta.[99] El númberu d'afecciones nes que podría tar implicáu esti xen ta n'aumentu. rellacionóse-y cola enfermedá de Moyamoya, y paez ser que delles mutaciones en heterocigosis podríen conferir predisposición a munches patoloxíes vasculares, como'l aneurisma d'aorta torácica y la cardiopatía isquémica.[100] La α actina de músculu llisu tamién ye un interesante marcador pa evaluar la progresión de la cirrosis hepática.[101]
De músculu cardiacu
ACTC1 ye'l xen que codifica la isoforma de la α actina presente nel músculu cardiacu. Secuenciar por primer vegada por Hamada y collaboradores en 1982, reparándose que taba atayáu por cinco intrones.[102] Foi'l primer xen de los seis onde s'atoparon apanfilos implicaos en procesos patolóxicos.[103]
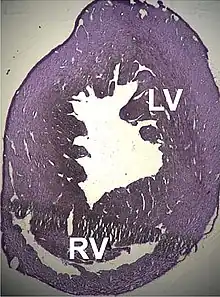
Describiéronse dellos trestornos estructurales que traen una disfunción cardiaca acomuñaos a mutaciones puntuales nesti xen, como miocardiopatía dilatada tipu 1R y la miocardiopatía hipertrófica tipu 11. Apocayá viose que dellos defectos atriales septales tamién podríen tar rellacionaos.[105][106]
Nel casu de la cardiomiopatía dilatada, estudiáronse dos casos nos qu'en dambos se produz una sustitución n'aminoácidos bien calteníos pertenecientes a los dominios que se xunen a los discos Z y entrepolaos, tou lo cual lleva a la hipótesis de que la dilatación producir por un defectu de trasmisión de la fuercia contráctil nos miocitos.[103][28]
Les alteraciones de ACTC1 son responsables de menos del 5% de les cardiomiopatías hipertrófiques.[107] Demostráronse tamién la esistencia de delles mutaciones puntuales:[108]
- Mutación Y101K: cambeos de carga neta y formación d'enllaz electrostático débil na posición d'unión de la actomiosina.
- P166A: zona d'interaición ente monómeros d'actina.
- A333P: zona d'interaición actina-miosina.
La patoxénesis paez obedecer a un mecanismu compensatoriu: les proteínes mutantes actuaríen como "tóxicu" con un efeutu dominante, menguando la capacidá de contraición con un rendimientu mecánicu anormal, de cuenta que la hipertrofia, que suel ser tardida, sería consecuencia d'una respuesta normal del músculu cardiacu al estrés.[109]
Apocayá atopáronse mutaciones de ACTC1 implicaes n'otros dos procesos patolóxicos: la miocardiopatía restrictiva idiopática infantil,[110] y el miocardiu ventricular esquierdu non compactu.[111]
D'actines citoplasmátiques
ACTB ye un locus bien complexu. Esisten ensame de pseudogenes partíos en tol xenoma, y la so secuencia contién seis exones que pueden dar llugar hasta 21 transcritos distintos por splicing alternativu, conocíos como actines β. En congruencia con esta complexidá, tamién los sos productos tienen llocalizaciones y formen parte de procesos bien distintos (citoesqueleto, complexu NuA4 histona-aciltransferasa, nucleu celular) y por lo mesmo tamién se-y acomuñó al mecanismu de gran cantidá de procesos patolóxicos (carcinomes, distonía xuvenil, mecanismos d'infeiciones, malformaciones nel sistema nerviosu y invasividad de neoplasmas, ente otres).[112] Atopóse una nueva forma d'actina, la actina kappa, que paez sustituyir a la actina β en procesos tumorales.[113]

Hasta'l momentu pudiéronse detectar tres proceso patolóxicos que se deben a una alteración direuta de la secuencia d'un xen:
- El hemangiopericitoma con translocación t(7;12)(p22;q13) ye una afección rara, na que se produz una fusión por translocación del xen ACTB sobre GLI1 nel cromosoma 12.[115]
- La distonía de debú xuvenil ye una enfermedá dexenerativa rara, con afectación sistémica del sistema nerviosu central, y especialmente d'árees neocorticales y talámicas, onde pueden apreciase un tipu d'inclusiones eosinofílicas en forma de cayáu. Los individuos afeutaos presenten un fenotipu con malformaciones na llinia media, perda auditiva sensorial y distonía. Deber a una mutación puntual que camuda l'aminoácidu arginina en posición 183 por un triptófanu. Esto alteria la interaición de la actina col sistema ADF/cofilina, que regula la dinámica de formación del citoesqueleto neuronal.[116]
- Atopóse una mutación puntual con calter dominante que produz disfunción de los neutrófilos y infeiciones recurrentes. Paez ser que la mutación modifica'l dominiu d'unión cola profilina y otres proteínes reguladores. L'afinidá pola profilina n'este apanfilo ta bien amenorgada.[117]
ACTG1 ye'l locus que codifica la proteína de la γ actina citosólica responsable de la formación de microfilamentos del citoesqueleto. Contién 6 exones, dando llugar a 22 mRNAs distintos, lo cual produz 4 isoformes completes, posiblemente espresaes d'una forma dependiente de texíu. Tamién tien dos promotor del ADN promotores alternativos.[118] Viose que les secuencies traducíes d'esti locus y el de la β actina son más asemeyaes de lo esperao, suxuriendo una secuencia ancestral común que sufrió duplicación y conversión xénica.[119]
Dende'l puntu de vista patolóxicu, foi acomuñáu a procesos como l'amiloidosis, la retinitis pigmentosa, mecanismos d'infeición, enfermedaes renales y diverses perdes auditives conxénites.[118]
Rellacionaes con seis mutaciones puntuales autosómico-dominantes na secuencia, atopamos diverses formes de perdes d'audición, cuantimás la sensorineural tipu 20/26. Paez qu'afecten de forma específica a los estereocilios de les célules ciliaes del órganu de Corti. La β actina ye la proteína más abondoso nos texíos humanos, pero non asina nes célules ciliaes, lo qu'esplicaría la llocalización de la patoloxía. Per otra parte, paez que la mayor parte d'estes mutaciones afecten a zones d'unión con otres proteínes, cuantimás la actomiosina.[26] Dellos esperimentos suxuren que'l mecanismu patogénico d'esti tipu de sordera deber a que la actina F de los mutantes sería más sensible de lo habitual a la cofilina.[120]
Per otra parte, anque nun se tien constancia de nengún casu, sábese que la γ actina tamién s'espresa nel músculu esqueléticu, y anque en cantidaes bien pequeñes, los modelos animales amosaron que la so ausencia podría dar llugar a miopatíes.[121]
Otros mecanismos patolóxicos
Dellos axentes infeiciosos utilicen la actina, especialmente la citoplasmática, na so ciclu de vida. En bacteries básicamente esisten dos formes:
- Listeria monocytogenes, delles especies de Rickettsia, Shigella flexneri y otros xermes intracelulares escapen de les vacuolas fagocíticas por aciu el recubrimientu con una cápsula curtia de filamentos d'actina. Nel casu de L. monocytogenes y S. flexneri, xeneren a partir d'ellos un cercu en forma de "cola de cometa" que dexa la so movilidá. Esisten llixeres diferencies nel mecanismu molecular de polimerización de la cola en cometa» dependiendo de la especie bacteria. Pueden reparase distintes velocidaes de desplazamientu, por casu, con un máximu pa Listeria y Shigella.[122] Munchos esperimentos ensayaron esti mecanismu in vitro. Estos amuesen que nun s'emplega nenguna proteína motor tipu miosina, y paez que la propulsión adquirir pola presión exercida pola polimerización que tien llugar cerca de la paré del microorganismu, que primeramente s'arrodió de ABP's mesmes de la célula hospedadora, que na so configuración mínima tratar del complexu Arp2/3, proteínes Ena-VASP, cofilina, una proteína taponante y promotores de la nucleación, como'l complexu de la vinculina. Por aciu estos movimientos formen protrusiones qu'algamen les célules vecines, infectándoles de la mesma, de cuenta que'l sistema inmunolóxicu namái puede combatir la infeición por aciu la inmunidá celular. La ruta del movimientu podría ser debida al cambéu de la combadura y desramificación de los filamentos.[123] Otres especies, como Mycobacterium marinum y Burkholderia pseudomallei, tamién son capaces de polimerizar llocalmente la actina celular pa facilitar el so desplazamientu por aciu un mecanismu que pivota sobre'l complexu Arp2/3; entá más, el virus vacunal o Vaccinia virus tamién emplega elementos del citoesqueleto d'actina pa la so diseminación.[124]
- Pseudomonas aeruginosa ye capaz de formar un biofilm proteutor col qu'escapa de les defenses del organismu, cuantimás de los neutrófilos y de los antibióticos, emplegando ADN y filamentos d'actina del hospedador.[125]
Amás del exemplu citáu enantes, nos pasos iniciales de la internalización de dalgunos virus, notablemente'l VIH, aguíyase la polimerización de la actina, por casu inactivando la cofilina.[126]
Nos procesos d'invasión de les célules canceroses, les protrusiones basaes en actina desempeñen un papel entá non determináu.[127]
Evolución
El citoesqueleto eucariota amuesa dellos componentes de gran semeyanza a lo llargo de la escala filoxenética, especialmente la actina y la tubulina. Por casu, la proteína codificada pol xen ACTG2 d'humanos tien una equivalencia absoluta colos ortólogos presentes n'aguarón y mure, magar a nivel de nucleótidos la identidá mengua al 92 %.[128] Sicasí, sí esisten mayores diferencies colos equivalentes en procariotas (FtsZ y MreB), que, de la mesma, presenten una identidá de secuencia d'ente un 40−50% ente les distintes especies de bacteries y arquies. Dellos autores suxuren que la proteína ancestral que dio llugar al modelu básicu d'actina eucariota asemeyar a les proteínes del citoesqueleto bacterianu presentes anguaño.[129]

Dellos autores resalten que la actina, la tubulina y les histones, un tipu de proteínes implicaes na estabilización y regulación del ADN, presenten semeyances na so capacidá de xunir nucleótidos y nel so funcionamientu basáu nel aprovechamientu del movimientu browniano; entá más, suxuren que toos ellos podríen derivar d'un ancestru común.[130] Poro, los mecanismos evolutivos diversificaron la proteína ancestral nes variantes güei presentes, calteniendo, ente otres, les actines como molécules eficaces pa encetar procesos biolóxicos antiguos y esenciales, como la endocitosis.[131]
Equivalentes en bacteries
Magar les bacteries nun tienen un citoesqueleto comparable en complexidá al de los eucariotes, describiéronse proteínes d'alta semeyanza colos monómeros y polímeros d'actina. La proteína MreB de bacteries polimeriza en filamentos delgaos, non helicoidales y, raramente, n'estructures helicoidales asemeyaes a la actina F.[17] Entá más, la so estructura cristalina ye bien asemeyada a la actina G (tocantes a conformanza tridimensional), ya inclusive esisten equivalencies ente los protofilamentos de MreB y la actina F. El citoesqueleto bacterianu tamién tien ente los sos componentes les proteínes FtsZ, asemeyaes a la tubulina.[132]
Poro, les bacteries tienen un citoesqueleto con elementos homólogos a la actina (por casu, MreB, ParM, y MamK), magar la secuencia aminoacídica d'estes proteínes diverxe de los presentes en célules animales. Sicasí, MreB y ParM tienen una alta semeyanza estructura de les proteínes estructural cola actina eucariota. Los microfilamentos, altamente dinámicos, xeneraos por aciu agregamientu de MreB y ParM son esenciales pa la viabilidá celular y participen na morfogénesis de la célula, segregación del genóforo y polaridá celular. ParM, un homólogu de la actina codificado nun plásmido, intervién na xestión del ADN plasmídico.[133]
Aplicaciones
L'aprovechamientu de la actina nos llaboratorios de ciencia y teunoloxía deriven de la so participación como riel de motores moleculares como la miosina (yá nel músculu, yá fora d'él), y de la so presencia necesaria pal funcionamientu celular. Tocantes a la clínica, yá que delles variantes anómales de la actina tán rellacionaes cola apaición de patoloxíes, la so detección ye un criteriu diagnósticu.
- Nanoteunoloxía. Los sistemes actina-miosina actúen como motores moleculares que dexen el tresporte de visícules y orgánulos a lo llargo de la citoplasma. Esisten esperimentos qu'aprovechen esta capacidá dinámica inclusive in vitro, esto ye, en sistemes acelulares, polo que se postuló una aplicación nanoteunolóxica del sistema. La idea subxacente ye emplegar los microfilamentos como rieles sobre los cualos una o más proteínes motores esmúzanse tresportando una determinada carga; esto ye, definir un circuitu espacial pol que pueda tresportase de forma empobinada y más o menos controlada una determinada carga. Tocantes a aplicaciones xenerales, falar del tresporte empobináu de molécules pa llograr la so lliberación en llugares concretos, lo que dexaría al ensamblaxe de nanoestructuras de forma controlada.[134] Estes capacidaes podríen ser aplicaes en chips d'investigación como los lab-on-a-chip, en nanocomponentes mecánicos y en nanotransformadores d'enerxía mecánica n'eléctrica.[135]
- Control internu en téuniques de bioloxía molecular, como'l western blot y la PCR en tiempu real. Por cuenta de que la función de la actina ye necesaria pa la sobrevivencia celular, postulóse que la so cantidá ta tan controlada a nivel de producción celular que puede asumise que la so trescripción (esto ye, el grau d'espresión de les sos xenes) y traducción, (que ye la xeneración de proteína) ye práuticamente constante, independientemente de les condiciones esperimentales. Por esta razón, nos estudios de cuantificación de proteínes (western blot) y de transcritos (PCR en tiempu real) suel realizase amás de la cuantificación del xen d'interés la d'un xen de referencia, como la mentada actina. Estremando la cantidá del xen d'interés pola de la actina ye posible llograr una cantidá relativa comparable ente distintos esperimentos,[136] siempres y cuando la espresión d'esta postrera nun varie; cabo destacar que la actina non siempres presenta la estabilidá deseyada na so espresión.[137]
- Clínica. Dalgunos apanfilos de la actina son causantes de patoloxíes, polo que se desenvolvieron téuniques pa la so detección. Amás, la actina puede emplegase como marcador indireutu en patoloxía quirúrxica: ye posible emplegar variaciones en pauta de llocalización nos texíos como marcadores d'invasión de neoplasies, vasculitis y otros.[138] Tamién, por cuenta de la so rellación col aparatu contráctil muscular, l'atrofia provoca l'amenorgamientu de los sos niveles nel músculu esqueléticu, polo que puede emplegase como marcador d'esti fenómenu.[139]
- Teunoloxía de los alimentos. La determinación de la calidá de dellos alimentos procesaos, como los embutíos, pasa pola cuantificación del so conteníu en carne. Clásicamente utilizóse un métodu basáu na detección de la 3-metilhistidina n'hidrolizados d'estos productos, pos se trata d'un compuestu presente na actina y la cadena pesada de la miosina F (dambos componentes mayoritarios del músculu). La xeneración nel animal del compuestu deber a la metilación de residuos de histidina presentes en dambes proteínes.[140][141]
Ver tamién
- Proteínes d'unión a actina
- Arp2/3
- Falotoxina
- Filopodio
- Filamentu entemediu
- FtsZ
- Lamelipodio
Notes
- ↑ Nesti enllaz reparar un modelu de la prefoldina cola actina encaxada ente los "tentáculos" de los sos subunidades.
- ↑ Nel siguiente enllaz d'un trabayu de Jaime Martín-Benito y José María Valpuesta, del centru nacional de Bioteunoloxía del CSIC, puede apreciase la configuración en doble aniellu de la chaperonina CCT, según les sos subunidades.
Referencies
- ↑ «The evolution of compositionally and functionally distinct actin filaments». Journal of Cell Science 128 (11): páxs. 2009-19. 2015. doi:. PMID 25788699.
- ↑ «In search of the primordial actin filament.». Proc Natl Acad Sci U S A. 112 (30): páxs. 9150-1. 2015. doi:. PMID 26178194.
- 1 2 3 4 Marc Maillet Bioloxía celular (n'español). Publicáu por Elsevier España, 2002; páx.132. ISBN 84-458-1105-3
- 1 2 Alberts et al (2004). Bioloxía molecular de la célula. Barcelona: Omega. ISBN 54-282-1351-8.
- ↑ Halliburton, W.D. (1887). «On muscle plasma». J. Physio. (8).
- ↑ «Atomic model of the actin filament», Nature 347 (6288), doi:, http://www.nature.com/nature/journal/v347/n6288/abs/347044a0.html
- ↑ Szent-Gyorgyi, A. (1945) Studies on muscle. Acta Physiol Scandinav 9 (suplementu. 25)
- ↑ Straub, F.B. y Feuer, G. (1950). «Adenosinetriphosphate the functional group of actin.». Biochim.Biophys. Acta.. PMID 2673365.
- ↑ Bárány, M., Barron, J.T., Gu, L., y Bárány, K. (2001). «Exchange of the actin-bound nucleotide in intact arterial smooth muscle». J. Biol. Chem. 276 (51). PMID 11602582.
- 1 2 «Complete amino-acid sequence of actin of rabbit skeletal muscle», Proceedings of the National Academy of Sciences 70 (9), http://www.pnas.org/cgi/reprint/70/9/2687.pdf, consultáu'l 29 de xunu de 2009
- 1 2 Kabsch W, Mannherz HG, Suck D, Pai EF, Holmes KC. (1990). «Atomic structure of the actin:DNase I complex.». Nature 347 (6288). PMID 2395459.
- ↑ Holmes KC, Popp D, Gebhard W, Kabsch W (1990). «Atomic model of the actin filament». Nature 347 (6288). PMID 2395461.
- 1 2 3 4 Domínguez, R; Otterbein LR, Graceffa P (Julio 2001) (n'inglés). The crystal structure of uncomplexed actin in the ADP state. 293. páxs. 708-11. PMID 11474115.
- ↑ Oriol, C; Dubord, C y Landon, F (xineru 1977). «Crystallization of native striated-muscle actin» (n'inglés). FEBS Lett. 73 (1): páxs. 89-91. PMID 320040.
- ↑ Sawaya MR, Kudryashov DS, Pashkov I, Adisetiyo H, Reisler Y, Yeates TO. (2008). «Multiple crystal structures of actin dimers and their implications for interactions in the actin filament». Acta Crystallogr D Biol Crystallogr.. PMID 18391412.
- ↑ Narita; Takeda S, Yamashita A, Maéda Y. (payares 2006). «Structural basis of actin filament capping at the barbed-end: a cryo-electron microscopy study» (n'inglés). Embo J 25 (23): páxs. 5626-33. doi:. http://www.nature.com/emboj/journal/v25/n23/pdf/7601395a.pdf. Consultáu'l 27 de xunu de 2009.
- 1 2 3 4 5 6 Toshiro Oda, Mitsusada Iwasa, Tomoki Aihara, Yuichiro Maéda y Akihiro Narita (2009): The nature of the globular- to fibrous-actin transition. Nature 457, 441-445 (22 January 2009) | doi:10.1038/nature07685
- ↑ «Human actin xenes are single copy for alpha-skeletal and alpha-cardiac actin but multicopy for β- and γ-cytoskeletal xenes: 3' untranslated regions are isotype specific but are conserved in evolution», Molecular and Cellular Biology 3 (10), http://mcb.asm.org/cgi/content/abstract/3/10/1783
- 1 2 3 4 Lodish et al. (2005). Bioloxía celular y molecular. Buenos Aires: Médica Panamericana. ISBN 950-06-1974-3.
- ↑ Futoshi Hara, Kan Yamashiro, Naoki Nemoto, Yoshinori Ohta, Shin-ichi Yokobori, Takuo Yasunaga, Shin-ichi Hisanaga, and Akihiko Yamagishi. (2007): An Actin Homolog of the Archaeon Thermoplasma acidophilum That Retains the Ancient Characteristics of Eukaryotic Actin. Journal of Bacteriology, p. 2039-2045, Vol. 189, No. 5 doi:10.1128/JB.01454-06
- 1 2 «Crystal Structure of Monomeric Actin in the ATP State: STRUCTURAL BASIS OF NUCLEOTIDE-DEPENDENT ACTIN DYNAMICS», Journal of Biological Chemistry 278 (36), doi:, PMID 12813032, http://www.jbc.org/cgi/content/full/278/36/34172
- ↑ «Actin molecular structure and function», Curr Opin Cell Biol 5 (1), doi:, http://www.ncbi.nlm.nih.gov/pubmed/8448029
- ↑ «NCBI Conserved Domains: ATP binding site» (inglés). Consultáu'l 26 d'avientu de 2008.
- 1 2 Elzinga; Collins, JH (agostu 1975). «The primary structure of actin from rabbit skeletal muscle. Completion and analysis of the amino acid sequence» (n'inglés). J Biol Chem. 250 (15). http://www.ncbi.nlm.nih.gov/pubmed/1150665?dopt=Abstract.
- ↑ Elzinga; Collins JH, Kuehl WM, Adelsteiskeletal muscle. (n'inglés)Proc Natl Acad Sci U S A. 70 (9). http://web.archive.org/web/http://www.pubmedcentral.nih.gov/picrender.fcgi?artid=427084&blobtype=pdf.
- 1 2 3 4 5 6 7 8 Cristóbal G. Dos Remedios, Deepak Chhabra (2008). Actin-binding Proteins and Disease. Springer. ISBN 0-387-71747-1. Ver en Google books
- ↑ Rould; Wan Q, Joel PB, Lowey S, Trybus KM. (ochobre 2006). «Crystal structures of expressed non-polymerizable monomeric actin in the ADP and ATP states». J Biol chem 281 (42). doi:. http://www.jbc.org/cgi/content/full/281/42/31909?view=long&pmid=16920713.
- 1 2 Devlin, Thomas M (2004). «23», a+la+actina+residuu&source=bl&ots=YZZKRVRbRQ&sig=WqMv1TZlXS6hakU88tdHgNLfbt4&hl=es&ei=9yZHSrjVBuPTjAeF4aRk&sa=X&oi=book_result&ct=result&resnum=4 Bioquímica: Llibru de testu con aplicaciones clíniques, 4 (n'inglés), Reverte, páx. 1021. ISBN 8429172084..
- 1 2 3 Egelman; Reisler, Y (avientu 2007). «Actin structure and function: what we still do not understand». J Biol Chem 282 (50). doi:. http://www.jbc.org/cgi/content/full/282/50/36133.
- ↑ DA Begg, R Rodewald y LI Rebhun (1978): The visualization of actin filament polarity in thin sections. Evidence for the uniform polarity of membrane-associated filaments. The Journal of Cell Biology, Vol 79, 846-852.
- 1 2 3 Arthur C. Guyton, John Y. Hall Tratáu de fisioloxía médica (n'español). Publicáu por Elsevier España, 2007; páx.76. ISBN 84-8174-926-5
- 1 2 Simons, CT; Staes A, Rommelaere H, Ampe C, Lewis SA, Cowan NJ (febreru 2004). «Selective contribution of eukaryotic prefoldin subunits to actin and tubulin binding» (n'inglés). J Biol Chem. 279 (6): páxs. 4196-203. doi:. PMID 14634002. http://www.jbc.org/cgi/content/full/279/6/4196?view=long&pmid=14634002#REF21.
- ↑ Martín-Benito (avientu 2002). «Structure of eukaryotic prefoldin and of its complexes with unfolded actin and the cytosolic chaperonin CCT» (n'inglés). EMBO J 21 (23): páxs. 6377-86. doi:. PMID 12456645. http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=12456645.
- 1 2 Vandamme, D; Lambert Y, Waterschoot D, Cognard C, Vandekerckhove J, Ampe C, Constantin B, Rommelaere H. (xunetu 2009). «alpha-Skeletal muscle actin nemaline myopathy mutants cause cell death in cultured muscle cells» (n'inglés). Biochim Biophys Acta. 1793 (7): páxs. 1259-71. PMID 19393268.
- 1 2 Brackley; Grantham J (xineru 2009). «Activities of the chaperonin containing TCP-1 (CCT): implications for cell cycle progression and cytoskeletal organisation» (n'inglés). Cell Stress Chaperones 14 (1): páxs. 23-31. doi:. PMID 18595008. http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=18595008.
- 1 2 Stirling, PC; Cuéllar J, Alfaro GA, El Khadali F, Beh CT, Valpuesta JM, Melki R, Leroux MR (marzu 2006). «PhLP3 modulates CCT-mediated actin and tubulin folding via ternary complexes with substrates» (n'inglés). J Biol Chem 281 (11): páxs. 7012-21. doi:. PMID 16415341. http://www.jbc.org/cgi/content/full/281/11/7012?view=long&pmid=16415341.
- ↑ Hansen, WJ; Cowan NJ, Welch WJ. (abril 1999). «Prefoldin-nascent chain complexes in the folding of cytoskeletal proteins» (n'inglés). J Cell Biol. (265-7) (145): p. 2. PMID 10209023. http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=10209023.
- ↑ Neirynck; Waterschoot D, Vandekerckhove J, Ampe C, Rommelaere H (xineru 2006). «Actin interacts with CCT via discrete binding sites: a binding transition-release model for CCT-mediated actin folding». J Mol Biol. 355 (1): páxs. 124-38. PMID 16300788.
- 1 2 3 Vavylonis, D; Yang Q, O'Shaughnessy B. (xunu 2005). «Actin polymerization kinetics, cap structure, and fluctuations» (n'inglés). Proc Natl Acad Sci U S A. 102 (24): páxs. 8543-8. doi:. PMID 15939882. http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=15939882.
- ↑ «Electron Microscopic Particle Length of F-Actin Polymerized in vitro», Journal of Biochemistry 67 (3), http://jb.oxfordjournals.org/cgi/content/abstract/67/3/437
- ↑ Cooper, Geoffrey M.; Robert Y. Hausman (2007). «Chapter 12: The Cytoskeleton and Cell Movement», The cell: a molecular approach (n'inglés). ASM Press, Washington. ISBN 0878932194.
- ↑ «Tabla de les constantes d'asociación y disociación de les distintes especies de monómeros al filamentu d'actina, según la lliteratura científica.».
- ↑ «Implications of treadmilling for the stability and polarity of actin and tubulin polymers in vivo», The Journal of Cell Biology 86 (1), PMID 6893454, http://www.jcb.org/cgi/reprint/86/1/330.pdf
- ↑ Lewin, Benjamin (2006). Cells (Google books) (n'inglés), Jones & Bartlett Publishers. ISBN 9780763739058.
- ↑ Zhang, Duan-Sun; Valeria Piazza, Benjamin J. Perrin, Agnieszka K. Rzadzinska, J. Collin Poczatek, Mei Wang, Haydn M. Prosser, James M. Ervasti, David P. Corey, Claude P. Lechene (15 de xineru de 2012). «Multi-isotope imaging mass spectrometry reveals slow protein turnover in hair-cell stereocilia» (n'inglés). Nature 481 (7382): páxs. 520-524. doi:. http://www.nature.com/doifinder/10.1038/nature10745. Consultáu'l 27 de xineru de 2012.
- ↑ THE STRUCTURE OF CRYSTALLINE PROFILIN-BETA-ACTIN Protein Data Bank
- ↑ Domínguez, R (payares 2004). Actin-binding proteins--a unifying hypothesis.. 29. páxs. 572-8. PMID 15501675.
- ↑ «The control of actin nucleotide exchange by thymosin β 4 and profilin. A potential regulatory mechanism for actin polymerization in cells», Molecular Biology of the Cell 3 (9), http://www.molbiolcell.org/cgi/content/abstract/3/9/1015
- ↑ Witke, W., Podtelejnikov, A., Di Nardo, A., Sutherland, J., Gurniak, C., Dotti, C., and M. Mann (1998) In Mouse Brain Profilin I and Profilin II Associate With Regulators of the Endocytic Pathway and Actin Assembly. The EMBO Journal 17(4): 967-976 Plantía:Entrez Pubmed
- ↑ Carlsson L, Nyström -Y, Sundkvist I, Markey F, Lindberg O. (1977) Actin polymerizability is influenced by profilin, a low molecular weight protein in non-muscle cells. J. Mol. Biol. 115:465-483 Plantía:Entrez Pubmed
- ↑ Kiselar, J., Janmey, P., Almo, S., Chance, M. (2003). «Visualizing the Ca2+-dependent activation of gelsolin by using synchrotron footprinting». PNAS 100 (7): páxs. 3942-3947. doi:. PMID 12655044. http://www.pnas.org/cgi/content/full/100/7/3942.
- ↑ «Gelsolin and ADF/cofilin enhance the actin dynamics of motile cells», Proceedings of the National Academy of Sciences of the United States of America 97 (13), doi:, PMID 10860951, http://www.pnas.org/cgi/content/full/pnas;97/13/6936
- ↑ «Effects of CapZ, an actin-capping protein of muscle, on the polymerization of actin», Biochemistry 28 (21), doi:, http://pubs.acs.org/doi/abs/10.1021/bi00447a036
- ↑ «Tropomodulin caps the pointed ends of actin filaments», The Journal of Cell Biology 127 (6), doi:, PMID 7798317, http://www.jcb.org/cgi/reprint/127/6/1627.pdf
- ↑ Robinson RC, Turbedsky K, Kaiser DA, Marchand JB, Higgs HN, Choe S, Pollard TD. (2001) Crystal structure of Arp2/3 complex (n'inglés). Science 294(5547):1679-84.
- ↑ Mullins, R. D.; Pollard, T.D. (abril de 1999). «Structure and function of the Arp2/3 complex». Current Opinion in Structural Biology (Elsevier) 9 (2): páxs. 244-249. doi:. http://www.ingentaconnect.com/content/els/0959440x/1999/00000009/00000002/art80034?token=00431ce3643f038fde4775686f2357275c277b422c40465d483f2544446y7b6dea2. Consultáu'l 3 d'ochobre de 2007.
- ↑ Laura M Machesky; Kathleen L Gould (Febreru de 1999). «The Arp2/3 complex: a multifunctional actin organizer». Current Opinion in Cell Biology 11 (1): páxs. 117-121. doi:. http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6VRW-3W8SD3R-G&_user=10&_rdoc=1&_fmt=&_orig=search&_sort=d&view=c&_acct=C000050221&_version=1&_urlVersion=0&_userid=10&md5=c399y9d447df11d385cbefe9b44056d7.
- ↑ «Latrunculin alters the actin-monomer subunit interface to prevent polymerization», Nature Cell Biology 2 (6), doi:, http://www.era.lib.ed.ac.uk/retrieve/1796/Mclaughlin.pdf
- 1 2 «Effects of cytochalasin and phalloidin on actin», The Journal of Cell Biology 105 (4), doi:, http://www.jcb.org/cgi/reprint/105/4/1473.pdf
- ↑ «Disruption of actin microfilaments by cytochalasin D leads to activation of p53», FEBS Letters 430 (3), doi:, http://linkinghub.elsevier.com/retrieve/pii/S0014579398006929
- ↑ «Cytochalasin D does not inhibit gravitropism in roots», American Journal of Botany 84 (11), doi:, http://www.amjbot.org/cgi/reprint/84/11/1530.pdf
- 1 2 «Actin and myosin as transcription factors», Current opinion in genetics & development 16 (2), doi:, http://web.archive.org/web/http://linkinghub.elsevier.com/retrieve/pii/S0959437X06000232
- ↑ «Phospholipase D activity is essential for actin localization and actin-based motility in Dictyostelium», Biochemical Journal 389 (Pt 1), doi:, http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1184553
- ↑ Randall, D.; Burggren, W. et French, K.. Eckert Fisioloxía animal, 4ª. ISBN 84-486-0200-5.
- ↑ Pujol-Moix (2001). Trombocitopenias, 2da, Elsevier, España, páx. 25. ISBN 8481745952.
- 1 2 3 Paniagua, R.; Nistal, M.; Sesma, P.; Álvarez-Uría, M.; Fraile, B.; Anadón, R. y José Sáez, F. (2002). Citoloxía ya histoloxía vexetal y animal. McGraw-Hill Interamericana d'España, S.A.U.. ISBN 84-486-0436-9.
- 1 2 «The Yeast Actin Cytoskeleton: from Cellular Function to Biochemical Mechanism», Microbiology and Molecular Biology Reviews 70 (3), doi:, PMID 16959963, http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1594590
- ↑ «Isovariant Dynamics Expand and Buffer the Responses of Complex Systems: The Diverse Plant Actin Gene Family», The Plant Cell Online 11 (6), http://www.plantcell.org/cgi/content/full/11/6/995
- ↑ Entrada PDBe pa 1unc. EBI.
- 1 2 «Actin microfilament dynamics and actin side-binding proteins in plants», Current Opinion in Plant Biology 10 (6), doi:, http://lmbiologia.campusnet.unito.it/didattica/att/a29a.1275.file.pdf (enllaz rotu disponible n'Internet Archive; ver l'historial y la última versión).
- ↑ «AtFim1 is an actin filament crosslinking protein from Arabidopsis thaliana», The Plant Journal 24 (5), doi:
- ↑ Antonio Bayés de Lluna, VV Staff, José López-Sendón, Fause Attie, Eduardo Allegría Ezquerra Cardioloxía Clínica (n'español). Publicáu por Elsevier España, 2002; páx.páx.19. ISBN 84-458-1179-7
- 1 2 John W. Baynes, Marek H. Dominiczak Bioquímica médica (n'español). Publicáu por Elsevier España, 2007; páx.268. ISBN 84-8174-866-8
- ↑ Randall, D.; Burggren, W. et French, K.. Eckert Fisioloxía animal, 4ª. ISBN 84-486-0200-5.
- ↑ «Alpha-actinin localization in the cleavage furrow during cytokinesis», The Journal of Cell Biology 79 (1), doi:, PMID 359574, http://www.jcb.org/cgi/reprint/79/1/268.pdf
- ↑ «Actin dynamics in the contractile ring during cytokinesis in fission yeast», Nature 419 (6902), http://adsabs.harvard.edu/abs/2002Natur.419...82P
- ↑ «Actin cleavage by CPP-32/apopain during the development of apoptosis», Oncogene 14 (9), doi:, http://www.nature.com/onc/journal/v14/n9/abs/1200919a.html
- ↑ «Calpain and caspase: can you tell the difference?», Trends in Neurosciences 23 (1), doi:, http://www.mbi.ufl.edu/ctbis/wang/Wang71TINS2000.pdf (enllaz rotu disponible n'Internet Archive; ver l'historial y la última versión).
- ↑ «Calpain inhibitors, but not caspase inhibitors, prevent actin proteolysis and DNA fragmentation during apoptosis», Journal of Cell Science 111 (6), http://jcs.biologists.org/cgi/reprint/111/6/713.pdf
- ↑ «SAPK2/p38-dependent F-Actin Reorganization Regulates Early Membrane Blebbing during Stress-induced Apoptosis», The Journal of Cell Biology 143 (5), http://www.jcb.org/cgi/content/full/143/5/1361
- ↑ «Quantitative analysis of cadherin-catenin-actin reorganization during development of cell-cell adhesion», The Journal of Cell Biology 135 (6), doi:, PMID 8991100, http://www.jcb.org/cgi/reprint/135/6/1899.pdf
- ↑ «Redundancy in the microfilament system: abnormal development of Dictyostelium cells lacking two F-actin cross-linking proteins», Cell 68 (1), http://www.ncbi.nlm.nih.gov/pubmed/1732064
- ↑ «Actin Plays a Role in Both Changes in Cell Shape and Gene- Expression Associated with Schwann Cell Myelination», Journal of Neuroscience 17 (1), http://www.jneurosci.org/cgi/content/full/17/1/241
- ↑ «Role of Actin Cytoskeletal Dynamics in Activation of the Cyclic AMP Pathway and HWP1 Gene Expression in Candida albicans», Eukaryotic Cell 6 (10), doi:, PMID 17715368, http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2043390
- ↑ «Novel Actin-Like Proteins T-ACTIN 1 and T-ACTIN 2 Are Differentially Expressed in the Cytoplasm and Nucleus of Mouse Haploid Germ Cells», Biology of Reproduction 69 (2), doi:, PMID 12672658, http://www.biolreprod.org/cgi/reprint/69/2/475.pdf (enllaz rotu disponible n'Internet Archive; ver l'historial y la última versión).
- ↑ «Epigenetic effects on yeast transcription caused by mutations in an actin-related protein present in the nucleus», Xenes & Development 10 (5), doi:, http://www.genesdev.org/cgi/reprint/10/5/604.pdf
- ↑ «Dynamic length regulation of sensory stereocilia», Seminars in Cell and Developmental Biology 19 (6), http://linkinghub.elsevier.com/retrieve/pii/S1084952108000451, consultáu'l 20 de xunu de 2009
- ↑ «An actin molecular treadmill and myosins maintain stereocilia functional architecture and self-», Journal of Cell Biology 164 (6), http://jcb.rupress.org/cgi/content/full/164/6/887, consultáu'l 20 de xunu de 2009
- ↑ El so AI, Wiltshire T, Batalov S, Lapp H, Ching KA, Block D, Zhang J, Soden R, Hayakawa M, Kreiman G, Cooke MP, Walker JR, Hogenesch JB (2004). «A gene atlas of the mouse and human protein-encoding transcriptomes». Proc Natl Acad Sci U S A. 101 (16). PMID 15075390. http://www.pnas.org/content/101/16/6062.full.
- 1 2 «Uniprot ACTA1» (inglés). Consultáu'l 26 d'avientu de 2008.
- 1 2 Friederike S Bathe, Heidi Rommelaere y Laura M Machesky (2007): Phenotypes of Myopathy-related Actin Mutants in differentiated C2C12 Myotubes. BMC Cell Biology, 8:2. doi:10.1186/1471-2121-8-2
- ↑ Kaindl AM, Rüschendorf F, Krause S, Goebel HH, Koehler K, Becker C, Pongratz D, Müller-Höcker J, Nürnberg P, Stoltenburg-Didinger G, Lochmüller H, Huebner A. (2004). «Missense mutations of ACTA1 cause dominant congenital myopathy with cores». J Med Genet. 41 (11). PMID 15520409.
- ↑ Sparrow, JC; Nowak KJ, Durling HJ, Beggs AH, Wallgren-Pettersson C, Romero N, Nonaka I, Laing NG. (setiembre 2003). «Muscle disease caused by mutations in the skeletal muscle alpha-actin gene (ACTA1)». Neuromuscul Disord. 13 (8): páxs. 519-31. PMID 12921789.
- ↑ North, Kathryn (2002). «Nemaline Myopathy». Gene Reviews. PMID. http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=gene&part=nem.
- ↑ Ilkovski B, Nowak KJ, Domazetovska A, Maxwell AL, Clement S, Davies KE, Laing NG, North KN, Cooper ST. (2004). «Evidence for a dominant-negative effect in ACTA1 nemaline myopathy caused by abnormal folding, aggregation and altered polymerization of mutant actin isoforms». Hum. Mol. Genet. 13 (16). PMID 15198992. http://hmg.oxfordjournals.org/cgi/content/full/13/16/1727.
- ↑ Clarke NF, Ilkovski B, Cooper S, Valova VA, Robinson PJ, Nonaka I, Feng JJ, Marston S, North K (2007). «The pathogenesis of ACTA1-related congenital fiber type disproportion». Ann Neurol 61 (6). PMID 17387733.
- ↑ «Structure, chromosome location, and expression of the human smooth muscle (enteric type) γ-actin gene: evolution of six human actin gen». Mol Cell Biol. 11 (6). 1991. PMID 1710027. http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=1710027.
- ↑ Watson MB, Lind MJ, Smith L, Drew PJ, Cawkwell L. (2007). «Expression microarray analysis reveals xenes associated with in vitro resistance to cisplatin in a cell line model». Acta Oncol. 46 (5). PMID 17562441.
- ↑ Guo DC, Pannu H, Tran-Fadulu V, Papke CL, Yu RK, Avidan N, Bourgeois S, Estrera AL, Safi HJ, Sparks Y, Amor D, Ades L, McConnell V, Willoughby CE, Güelu D, Willing M, Lewis RA, Kim DH, Scherer S, Tung PP, Ahn C, Buja LM, Raman CS, Shete SS, Milewicz DM. (2007). «Mutations in smooth muscle alpha-actin (ACTA2) lead to thoracic aortic aneurysms and dissections». Nature Genet. 39 (12). PMID 17994018.
- ↑ Guo; Papke CL, Tran-Fadulu V, Regalado YE, Avidan N, Johnson RJ, Kim DH, Pannu H, Willing MC, Sparks Y, Pyeritz RE, Singh MN, Dalman RL, Grotta JC, Marian AJ, Boerwinkle EA, Frazier LQ, LeMaire SA, Coselli JS, Estrera AL, Safi HJ, Veeraraghavan S, Muzny DM, Wheeler DA, Willerson JT, Yu RK, Shete SS, Scherer SE, Raman CS, Buja LM, Milewicz DM. (mayu 2009). «Mutations in smooth muscle alpha-actin (ACTA2) cause coronary artery disease, stroke, and Moyamoya disease, along with thoracic aortic disease» (n'inglés). Am J Human Genet 84 (5): páxs. 617-27. PMID 19409525.
- ↑ Akpolat N, Yahsi S, Godekmerdan A, Yalniz M, Demirbag K (2005). «The value of alpha-SMA in the evaluation of hepatic fibrosis severity in hepatitis B infection and cirrhosis development: a histopathological and immunohistochemical study». Histopathology 47 (3). PMID 16115228.
- ↑ Hamada H, Petrino MG, Kakunaga T. (1982). «Molecular structure and evolutionary origin of human cardiac muscle actin gene». Proc Natl Acad Sci U S A. 79 (19). PMID 6310553. http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=6310553.
- 1 2 Olson TM, Michels VV, Thibodeau SN, Tai YS, Keating MT. (1998). «Actin mutations in dilated cardiomyopathy, a heritable form of heart failure.». Science 280 (5364). PMID 9563954.
- ↑ Xia X-G, Zhou H, Samper Y, Melov S, Xu Z (2006) Pol II–Expressed shRNA Knocks Down Sod2 Gene Expression and Causes Phenotypes of the Gene Knockout in Mice. PLoS Genet 2(1): y10. doi:10.1371/journal.pgen.0020010
- ↑ «OMIM 102540» (inglés). Consultáu'l 27 d'avientu de 2008. (enllaz rotu disponible n'Internet Archive; ver l'historial y la última versión).
- ↑ Matsson H, Eason J, Bookwalter CS, Klar J, Gustavsson P, Sunnegårdh J, Enell H, Jonzon A, Vikkula M, Gutiérrez I, Granados-Riveron J, Pope M, Bu'Lock F, Cox J, Robinson TE, Song F, Brook DJ, Marston S, Trybus KM, Dahl N. (2008). «Alpha-cardiac actin mutations produz atrial septal defects.». Hum Mol Genet. 17 (2). PMID 17947298.
- ↑ Kabaeva, Kabaeva,Z. «Genetic analysis in hypertrophic cardiomyopathy: missense mutations in the ventricular myosin regulatory light chain gene» (inglés). Universidá Humboldt de Berlín. Consultáu'l 27 d'avientu de 2008.
- ↑ Olson TM, Doan TP, Kishimoto NY, Whitby FG, Ackerman MJ, Fananapazir L. Inherited and de novo mutations in the cardiac actin gene cause hypertrophic cardiomyopathy (n'inglés). J Mol Cell Cardiol 2000; 32:1687-1694.
- ↑ Ramírez, Carlos Darío. «Cardiomiopatía hipertrófica familiar: Xenes, mutaciones y modelos animales. Revisión.». Invest. clín, mar. 2004, vol.45, non.1, p.69-100. Consultáu'l 15 de xunetu de 2009.
- ↑ Kaski JP, Syrris P, Burch M, Tomé-Esteban MT, Fenton M, Christiansen M, Andersen PS, Sebire N, Ashworth M, Deanfield JE, McKenna WJ, Elliott PM. (2008). «Idiopathic restrictive cardiomyopathy in children is caused by mutations in cardiac sarcomere protein gen». Heart 94 (11). PMID 18467357.
- ↑ Klaassen S, Probst S, Oechslin Y, Gerull B, Krings G, Schuler P, Greutmann M, Hürlimann D, Yegitbasi M, Pons L, Gramlich M, Drenckhahn JD, Heuser A, Berger F, Jenni R, Thierfelder L. (2008). «Mutations in sarcomere protein genes in left ventricular noncompaction». Circulation 117 (22). PMID 1850600.
- ↑ «ACEVieW:Homo sapiens complex locus ACTB, encoding actin, β.» (inglés). Consultáu'l 28 de xunu de 2008.
- ↑ Chang KW, Yang PY, Lai HY, Yeh TS, Chen TC, Yeh CT. (2006). «Identification of a novel actin isoform in hepatocellular carcinoma». Hepatol Res. 36 (1). PMID 16824795.
- ↑ Kristy L Williams, Masuma Rahimtula yKaren M Mearow. Hsp27 and axonal growth in adult sensory neurons in vitro BMC Neuroscience 2005, 6:24doi:10.1186/1471-2202-6-24
- ↑ «Pericytoma with t(7;12)». Atlas of Genetics and Cytogenetics in Oncology and Haematology. Consultáu'l 28 d'avientu de 2008.
- ↑ Procaccio V, Salazar G, Ono S, Styers ML, Gearing M, Dávila A, Jiménez R, Xuncos J, Gutekunst CA, Meroni G, Fontanella B, Sontag Y, Sontag JM, Faundez V, Wainer BH. (2006). «A mutation of β -actin that alters depolymerization dynamics is associated with autosomal dominant developmental malformations, deafness, and dystonia». Am J Hum Genet 78 (6). PMID 16685646. http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=16685646#RF21.
- ↑ Nunoi H, Yamazaki T, Tsuchiya H, Kato S, Malech HL, Matsuda I, Kanegasaki S. (1999). «A heterozygous mutation of β-actin associated with neutrophil dysfunction and recurrent infection». Proc Natl Acad Sci U S A. 96 (15). PMID 10411937. http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=10411937.
- 1 2 «ACEVIEW ACTG1» (inglés). Consultáu'l 29 d'avientu de 2008.
- ↑ Erba HP, Gunning P, Kedes L. (1986). «Nucleotide sequence of the human γ cytoskeletal actin mRNA: anomalous evolution of vertebrate non-muscle actin xenes.». Nucleic Acids Res. 14 (13). PMID 373740. http://web.archive.org/web/http://www.pubmedcentral.nih.gov/pagerender.fcgi?artid=311540&pageindex=2#page.
- ↑ Bryan, KE; Rubenstein, PA (xunetu 2009). «Allele-specific Effects of Human Deafness {γ}-Actin Mutations (DFNA20/26) on the Actin/Cofilin Interaction» (n'inglés). J Biol Chem. 284 (27): páxs. 18260-9. PMID 19419963.
- ↑ Sonnemann KJ, Fitzsimons DP, Patel JR, Liu Y, Schneider MF, Moss RL, Ervasti JM. (2006). «Cytoplasmic γ-actin is not required for skeletal muscle development but its absence leads to a progressive myopathy.». Dev Cell 11 (3). PMID 16950128.
- ↑ «A comparative study of the actin-based motilities of the pathogenic bacteria Listeria monocytogenes, Shigella flexneriand Rickettsia conorii», Journal of Cell Science 112 (11), https://journals.biologists.com/jcs/article/112/11/1697/25636/A-comparative-study-of-the-actin-based-motilities
- ↑ Lambrechts, A; Gevaert K, Cossart P, Vandekerckhove J, Van Troys M. (mayu 2008). «Listeria comet tails: the actin-based motility machinery at work» (n'inglés). Trends Cell Biol. 18 (5): páxs. 220-7. PMID 18396046.
- ↑ «Actin-based motility of intracellular pathogens», Current Opinion in Microbiology 8 (1), http://linkinghub.elsevier.com/retrieve/pii/S1369527404001675
- ↑ Parks, QM; Young RL, Poch KR, Malcolm KC, Vasil ML, Nick JA. (abril 2009). «Neutrophil enhancement of Pseudomonas aeruginosa biofilm development: human F-actin and DNA as targets for therapy». J med microbiol: páxs. 492-502 pmid=19273646. doi:. http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=19273646.
- ↑ Liu; Belkina NV, Shaw S. (abril 2009). «HIV infection of T cells: actin-in and actin-out» (n'inglés). Sci Signal. 2. PMID 19366992.
- ↑ Machesky LM; Tang HR (xunetu 2009). «Actin-Based Protrusions: Promoters or Inhibitors of Cancer Invasion?». Cancer Cell. 16 (1): páxs. 5-7. PMID 19573806.
- ↑ «The nucleotide sequence of a human smooth muscle (enteric type) γ-actin cDNA.», Molecular and Cellular Biology 11 (6), http://web.archive.org/web/http://www.pubmedcentral.nih.gov/picrender.fcgi?artid=360182
- ↑ «Evolution of the cytoskeleton», BioEssays: news and reviews in molecular, cellular and developmental biology 29 (7), http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2630885
- ↑ «Are histones, tubulin, and actin derived from a common ancestral protein?», Protoplasma 233, doi:
- ↑ «Actin and endocytosis: mechanisms and phylogeny», Current Opinion in Cell Biology 21, doi:, http://web.archive.org/web/http://linkinghub.elsevier.com/retrieve/pii/S0955067409000076
- ↑ «Prokaryotic origin of the actin cytoskeleton», Nature 413, doi:, http://web.archive.org/web/http://www2.mrc-lmb.cam.ac.uk/ss/Amos_L/group/PDF/MreB%20Nature.pdf
- ↑ «The Bacterial Actin-Like Cytoskeleton», Microbiology and Molecular Biology Reviews 70 (4), doi:, PMID 17158703, http://web.archive.org/web/http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1698507
- ↑ «Light-controlled molecular shuttles made from motor proteins carrying cargu on engineered surfaces», Nano Letters 1 (5), doi:
- ↑ «Actin-Based Molecular Motors for Cargu Transportation in Nanotechnology—Potentials and Challenges», IEEE Transactions on Advanced Packaging 28 (4), http://ieeexplore.ieee.org/xpls/abs_all.jsp?arnumber=1528636
- ↑ Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, Speleman F (2002) Accurate normalisation of real-time quantitative RT-PCR data by geometric averaging of multiple internal control gen. Xen. Biol. 3: 1–12.
- ↑ «ß-Actin», Molecular and cellular Probes 15 (5), http://web.archive.org/web/http://linkinghub.elsevier.com/retrieve/pii/S0890850801903767, consultáu'l 7 de xunetu de 2009
- ↑ «Immunohistochemical localization of actin: Applications in surgical pathology.», The American Journal of Surgical Pathology 5 (1), http://www.ajsp.com/pt/re/ajsp/abstract.00000478-198101000-00013.htm, consultáu'l 19 d'abril de 2009
- ↑ «Atrophy responses to muscle inactivity. II. Molecular markers of protein deficits», Journal of Applied Physiology 95 (2), http://jap.physiology.org/cgi/content/full/95/2/791, consultáu'l 19 de marzu de 2009
- ↑ «Current advances in proteomic analysis and its use for the resolution of poultry meat quality», World's Poultry Science Journal 62 (01), http://www.animalscience.com/uploads/additionalFiles/QualityOfPoultryMeat/27.pdf, consultáu'l 7 de xunetu de 2009
- ↑ M.L., «Methods and Instruments in Applied Food Analysis», Handbook of Food Analysis., http://orton.catie.ac.cr/cgi-bin/wxis.exe/?IsisScript=BFHIA.xis, consultáu'l 7 de xunetu de 2009
Enllaces esternos
 Wikimedia Commons acueye conteníu multimedia sobre Actina.
Wikimedia Commons acueye conteníu multimedia sobre Actina.
- Videu de la polimerización de la actina (n'inglés)
- Videu de la interaición actina-miosina (n'inglés)
- MMDB ID 50538 Modelu molecular de G-actina xunida a ATP por Domínguez.
- MMDB ID 69126 Modelu molecular de F actina por T. Oda