نظرية كانون- بارد
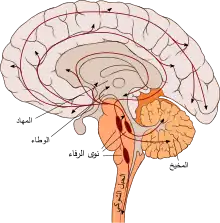
المفاهيم الرئيسية لنظرية كانون- بارد (بالإنجليزية: Cannon–Bard theory) هي أن التعبير الوجداني (أو الانفعالي أو الشعوري أو العاطفي) ينتج عن وظيفة البنى الوطائية (تحت المهادية)، والمشاعر الوجدانية تنتج عن تنبيه المهاد الظهراني. تتصف التغيرات الفسيولوجية والإحساس الشخصي بشعور ما استجابةً لمنبهات معينة بأنها منفصلةً ومستقلة؛ إذ لا يجب أن تحدث الإثارة قبل الانفعال، وبالتالي تلعب المنطقة المهادية دورًا رئيسيًا في نظرية الانفعال هذه. وبناءً على ذلك، سُميّت هذه النظرية أيضًا بالنظرية المهادية للانفعال.[1]
منشأ النظرية
كان والتر برادفورد كانون (1871 - 1945) عالم فسيولوجيا في جامعة هارفارد، وربما كانت أطروحته الكلاسيكية عن الاستتباب أفضل ما عُرف به.[2] كان فيليب بارد (1898- 1977) طالب دكتوراه تحت يدي والتر كانون، وطوّرا معًا نموذجًا للانفعال أطلقا عليه اسم نظرية كانون- بارد.[2][3] كان كانون مختبِرًا ومُجرِّبًا اعتمد في دراساته على فسيولوجيا الحيوان. أظهر كانون وبارد من خلال هذه الدراسات دور الدماغ في توليد الاستجابات الفسيولوجية والمشاعر، وهو دور مهم في التفسير الذي قدماه للتجارب الانفعالية وكيفية إنتاجها.[2]
كانت نظرية جيمس-لانج هي نظرية الانفعال السائدة في زمن كانون. أدرك كانون أن اختبار هذه النظرية يتطلب فحص التعبير الانفعالي الذي لا يتطلب التغذية الراجعة الواردة الحشوية، وكان هذا ضروريًا لأن الصلة بين التغيرات الحشوية والتغذية الراجعة المطلوبة لتنبيه التظاهرات الدماغية لانفعال ما لن يبق موجودًا.[4] ومن أجل القيام بذلك، استخدم كانون أعصابًا واردةً مقطوعةً من الجذع الودي للجهاز العصبي الذاتي عند القطط. جمع كانون هذه النتائج التجريبية عام 1915 ثم نقحها ونشرها، واقترح أخيرًا نموذجه في الانفعال باعتباره منافسًا وبديلًا لنظرية الانفعال لجيمس لانج.[2]
تعتمد نظرية جيمس-لانج[5] في تفسيرها للتجارب الانفعالية الفريدة على تدفق الدفعات العصبية من المحيط لتكون تجارب شعورية فريدة؛ إذ افترض وليام جيمس أن هذه الدفعات تأتي من جميع أجزاء العضوية، بما فيها الجلد والعضلات والأحشاء. نُسب للأحشاء -حسب رأي جيمس- الدور الرئيسي في ذلك. تتكون هذه الأحشاء من عضلات ملساء وغدد. حدد كانون وأوضح خمس نقاط يختلف فيها مع مفهوم نظرية جيمس لانج حول اعتبار المراكز الوعائية المحركة الموجودة في الدماغ تفسيرًا للتجربة الانفعالية.[6][7]
النقطة الأولى: لا يؤدي الفصل التام بين الأحشاء والجهاز العصبي المركزي إلى تغيير السلوك الانفعالي
في إحدى التجارب، بقيت القطط على قيد الحياة وفي صحة جيدة بعد الإزالة التامة لجهازها العصبي الودي. أدت إزالة هذا الجهاز إلى إلغاء جميع ردود الفعل الخاضعة لسيطرة المراكز المُحركة للأوعية، وهي المناطق التي اقترحت نظرية جيمس لانج أنها مسؤولة عن التجارب الانفعالية. وُجد أن تخريب وظائف هذه المراكز كان له تأثيرٌ قليلٌ أو معدوم في الاستجابات الانفعالية عند الحيوان، إذ أظهرت القطط علامات غضب نموذجية كرد فعل على نباح الكلاب، وأظهرت الحيوانات أيضًا تعبيرات انفعالية كاملة في جميع الأعضاء التي لم تفقد صلتها بالدماغ.[2][6]
النقطة الثانية: تحدث نفس التغيرات الحشوية في حالات انفعالية مختلفة بشدة، وفي حالات أخرى غير انفعالية
يعمل الجهاز العصبي الودي على أنه وحدة متكاملة. وتحدث التغيرات الحشوية بسبب فعل الجهاز العصبي الودي، ومن ضمنها: زيادة معدل ضربات القلب، وتضيّق الشرينات، وتوسع القصيبات، وزيادة مستوى السكر في الدم، والتعرق، وتوسع حدقة العين، وانتصاب الشعر، وإفراز الأدرينالين.
يمكن رؤية هذه التغيرات الفسيولوجية بأقصى درجاتها تحت أي ظرف من الظروف. وتتضمن الحالات الانفعالية القابلة للتمييز: الخوف والغضب وحالات الحمى والاختناق والتعرض لدرجات الحرارة المنخفضة.
أوضح كانون أن ردود الفعل الحشوية كانت منتظمة جدًا على أن تشكل طريقةً لتمييز الانفعالات التي تملك نوعية جسدية متنوعة. وافترض أنه إذا كانت الانفعالات ناتجة عن الدفعات العصبية القادمة من الأحشاء، فيمكننا أن نتوقع الخوف والغضب والبرودة والاختناق والحمى، وهذا ليس واقعيًا.
النقطة الثالثة: الأحشاء هي بنى غير حساسة نسبيًا
كتب كانون عن وجود اعتقاد شائع وهو أن حساسية أعضاء الجسم تزداد كلما تغلغلنا عميقًا بداخلها؛ إلا أن هذا ليس واقعيًا. الأعصاب المنتشرة في الأحشاء هي ألياف عصبية حسية واردة بمقدار 1/10 فقط من مجمل الألياف العصبية الحسية الواردة.[8] على سبيل المثال: نحن لا ندرك عمليات الانقباض والاسترخاء أثناء حدوث العمليات الهضمية داخل الأحشاء، إذ لا يمكننا التكهن بهذه العمليات التي تتخطى وعينا الجسدي، حتى عندما تحدث تغيرات واضحة فيها.[6]
النقطة الرابعة: التغيرات الحشوية بطيئة جدًا لتشكل مصدرًا للمشاعر الانفعالية
كما ذكر سابقًا، الأحشاء مكونة من مجموعة من العضلات الملساء والغدد التي تكون عادةً ذات استجابة بطيئة. وُجد أن فترة الكمون للاستجابة الغلفانية النفسية (الاستجابة للمحفزات العاطفية) عند الرجال هي 3 ثوان تقريبًا.[9] ولوحظ أن فترة الكمون للاستجابات العاطفية لصور الرجال والنساء يمكنها أن تنتهي خلال 0.8 ثانية. أكدت نظرية جيمس لانج أن هذه الاستجابات العاطفية تنتج عن انعكاسات من الأحشاء. وأشار كانون إلى أن الوقت اللازم لانتقال الدفعات العصبية من الدماغ إلى المحيط والعودة إلى الدماغ مرة أخرى لا يمكن أن يحدث بسرعة كافية ليكون سببًا لمثل هذه الاستجابات العاطفية.
النقطة الخامسة: التحريض الاصطناعي للتغيرات الحشوية بما يناسب الانفعالات القوية لا يسبب حدوث هذه الانفعالات
عند حقن الأدرينالين، فإنه يُحرّض الاستجابات الفسيولوجية النموذجية لنشاط الجهاز العصبي الودي كما نوقش سابقًا (توسع القصيبات وتضيق الأوعية الدموية وزيادة مستوى السكر في الدم وما إلى ذلك). هذه هي التغيرات النموذجية للحالات الانفعالية الشديدة، لذلك إذا نتجت هذه التغيرات الحشوية اصطناعيًا بعد حقن الأدرينالين، يمكن عندها توقع الانفعالات التي تليها، كما هو موضح في نظرية الانفعال لجيمس لانج. عند انتهاء هذه التجربة، لم يواجه المشاركون أي حالات انفعالية مميزة. ومع ذلك، وجد أنه يمكن للاستجابة الانفعالية أن تتطور فقط عندما يُحقن الأدرينالين بعد مناقشة المرضى بخصوص أطفالهم المرضى أو آبائهم المتوفين. وهكذا يكون لحقن الأدرينالين تأثيرًا فعالًا، أي عندما يكون المزاج الانفعالي موجودًا مسبقًا عند المشاركين.[6]
انتقادات إضافية لنظرية جيمس-لانج
طرح ويليام جيمس أن هناك مراكز خاصة للعمليات الدماغية التي ترافق الانفعال أو أنها تحدث في المراكز الحركية والحسية الاعتيادية في القشرة المخية.[5] وأجاب كانون مفترضًا أنه قد لا تكون الأولى أو الثانية، وأنه ربما يكون هناك عمليات قشرية ومراكز خاصة ترافق الاستجابات الانفعالية، وشرح باختصار فكرتين تتعلقان بوجود مصدرين للعمليات الانفعالية في الدماغ.
ينتج التعبير الانفعالي عن عمل المراكز تحت القشرية.
لخّص كانون البحث الذي أنجزه العالِم بيشتيريف حول التعبير الانفعالي.[10] إذ طُرح في هذا البحث أن التعبير الانفعالي يجب أن يكون مستقلًا عن القشرة الدماغية لأنه لا يمكننا دائمًا ضبط التعبيرات الانفعالية أو التحكم بها (مثلًا الضحك بسبب الدغدغة)، لأن التغيرات الحشوية تحدث بشكل خارج عن سيطرتنا، ولأن هذه الاستجابات -التي لا يمكننا منعها أو التحكم بها- يمكن رؤيتها حالًا بعد الولادة، وذلك قبل تطور التنظيم القشري. علاوةً على ذلك، يمكن حدوث استجابات انفعالية صحيحة نتيجة منبهات مناسبة، وذلك بعد إزالة نصفي الكرة المخية من نماذج اختبار حيوانية. إلا أنه بعد إزالة المهاد البصري (السرير البصري) عند الحيوانات، لم تعد هذه التأثيرات الانفعالية موجودة، وبالتالي استُنتج أن هذه المنطقة تلعب دورًا رئيسيًا في التعبير الانفعالي.
في دعم أكبر للتأكيد على أن التعبير الانفعالي ينتج عن عمل المراكز تحت القشرية، قدم كانون وبريتون[11] بحثًا تجريبيًا إضافيًا آخر باستخدام القطط. إذ أُزيلت القشرة المخية عند القطط، وبعد فترة من التعافي أبدت -بطريقة عفوية- صفات سلوكية مميزة عند تعرضها للغضب الشديد. واستمرت هذه الاستجابة العنيفة الكاذبة (الاستشاطة الكاذبة) في الظهور بعد استئصال جميع المناطق الدماغية أمام الدماغ البيني.[7][12] وبمجرد إزالة الجزء الخلفي السفلي من المنطقة المهادية، خمدت هذه الاستشاطة الكاذبة التي ظهرت عند القطط. بناءً على هذا الاكتشاف، ظهر أن المهاد هو المنطقة التي تنطلق منها الدفعات العصبية التي تثير بدورها درجةً شديدةً من النشاط الانفعالي العضلي والحشوي، وذلك في غياب التحكم القشري. وبناءً عل هذه النتائج والملاحظات، أكد كانون أن المهاد البصري هو منطقة في الدماغ مسؤولة عن التنظيم العصبي للتعبيرات الانفعالية المختلفة.[6]
مراجع
- J.E. Roeckelein (2006). Elsevier's Dictionary of Psychological Theories. Elsevier. ص. 85. ISBN:9780080460642. مؤرشف من الأصل في 2020-02-09.
- Friedman، B.H. (2010). "Feelings and the body: The Jamesian perspective on autonomic specificity of emotion". Biological Psychology. ج. 84 ع. 3: 383–393. DOI:10.1016/j.biopsycho.2009.10.006. PMID:19879320. مؤرشف من الأصل في 2020-02-09.
- American Physiological Society. "Philip Bard". مؤرشف من الأصل في 2016-09-24. اطلع عليه بتاريخ 2012-03-11.
- McCarty، R. (2007). "Fight-or-Flight Response". Encyclopedia of Stress (ط. 2nd). New York: Academic Press. ص. 62–64. DOI:10.1016/B978-012373947-6.00160-4. ISBN:9780123739476.
- James، W.؛ C.G. Lange (1922). The emotions. Baltimore: Williams & Wilkins Co. مؤرشف من الأصل في 2020-02-09.
- Cannon، W.B. (1927). "The James-Lange theory of emotions: A critical examination and an alternative theory". The American Journal of Psychology. ج. 39 ع. 1/4: 106–124. DOI:10.2307/1415404. JSTOR:1415404.
- Dalgleish، T. (2004). "The emotional brain". Nature Reviews Neuroscience. ج. 5 ع. 7: 582–589. DOI:10.1038/nrn1432. PMID:15208700. مؤرشف من الأصل في 2020-02-09.
- Langley، J.N.؛ H.K. Anderson (1894). "The constituents of the hypogastric nerves". Journal of Physiology. ج. 17 ع. 3–4: 177–91. DOI:10.1113/jphysiol.1894.sp000526. PMC:1514563. PMID:16992211.
- Wells، F.L.؛ A. Forbes (1911). "On certain electrical processes in the human body and their relations to emotional reactions". ج. 2 ع. 16: 8.
{{استشهاد بدورية محكمة}}: الاستشهاد بدورية محكمة يطلب|دورية محكمة=(مساعدة) - Bechterev، W. (1887). "Die bedeutung der sehhugel auf grund von experimentellen und pathologischen daten". Virchows Archiv. ج. 110.
- Cannon، W.B.؛ S.W. Britton (1925). "Pseudoaffective medulliadrenal secretion". American Journal of Physiology. ج. 72 ع. 2: 283–294. DOI:10.1152/ajplegacy.1925.72.2.283.
- Vinken، P.J. (1969). Handbook of Clinical Neurology. Amsterdam: North-Holland Publishing Group. ص. 316.
 بوابة علم النفس
بوابة علم النفس بوابة العقل والدماغ
بوابة العقل والدماغ