مشط الأسنان
مشط الأسنان أو المشط السني (بالإنجليزية: Toothcomb)[1] هو تركيب سني يوجد في بعض الثدييات. وهو يتألف من الأسنان الأمامية المُرتبة بطريقة تسهل عملية التنظيف، وهي شبيهة بمشط الشعر. يوجد مشط الأسنان في الحيوانات الليمورية الرئيسة، التي تشمل الليمور واللوريس، وزبابة الأشجار والكولوجو وحيوان الوبر، بالإضافة إلى بعض الظباء الأفريقية. تطورت تلك التراكيب بشكل مُستقل في أنواع مُختلفة من الثدييات من خلال التطور المُتقارب، وتنوعت في كل من التركيب السني والبنية. يتكون المشط في مُعظم الثدييات من مجموعة أسنان تفصلهم مسافات صغيرة. يتكون مشط الأسنان في مُعظم الثدييات من القواطع فقط، في حين أنها تتكون في الحيوانات الليمورية من القواطع والأسنان النابية التي تميل إلى الأمام في مُقدمة الفك السفلي، ويتبعها قبطاحن أول على هيئة ناب. ويتخذ مشط الأسنان لدى الكولوجو وحيوان الوبر شكلا مُختلفا، حيث تكون القواطع الفردية مُسننة، مُزودة كل سِنة بالعديد من الشوكات.

يُستخدم مشط الأسنان عادةً في التنظيف. فعندما يلعق الحيوان الفراء ليجعله نظيفًا، يمرر مشط الأسنان خلال الفراء لتمشيطه. تُقطع الأخاديد أو التصدعات الصغيرة عادةً في الأسنان أثناء تنظيف الشعر، ويمكن رؤيتها على جوانب الأسنان عند عرضها باستخدام المجهر الفاحص للإلكترونات.[2] يتم الحفاظ على نظافة مشط الأسنان باستخدام اللسان، أو باستخدام منطقة تحت اللسان بالنسبة لليموريات. ولمشط الأسنان وظائف أخرى أيضًا، مثل الحصول على الغذاء ونزع اللحاء. ومن بين الليموريات، فإن ليمور الشوكة المُتوج والإندرية يمتلكان مشط أسنان أقوى لدعم تلك الوظائف الثانوية. وقد فقدت بعض الليمورات مثل الآياي مشط الأسنان بالكامل، وتم استبداله بأسنان خاصة أخرى.[3]
استخدم العلماء مشط الأسنان لدى الليموريات الرئيسة لتفسير تطور الليمور، وأقاربه. حيث كان هناك ظن أنهم قد تطوروا من رئيسيات الحيوانات أدبيات الشكل في العصر الإيوسيني أو في وقت أسبق. ولكن هناك فرضية شائعة تقول بأنهم قد تطوروا من رئيسيات أوروبية منقرضة ظهرت خلال العصر الإيوسيني. ولكن تُشير السجلات الحفرية إلى أنهم قد تطوروا من سلسلة نسب أقدم من ذلك هاجرت إلى أفريقيا خلال العصر الباليوسيني، من قبل 65 مليون سنة إلى ما قبل 55 مليون سنة، أو من المُحتمل أنها قد تطورت من فصيلة السيركامونيات من أدبيات الشكل المُنقرضة المُبكرة من آسيا. ومن المُمكن أن يكون الجنس المُنقرض من الرئيسيات المُتحجرات الأولية من بداية العصر الإيوسيني مثل الليمور الجبلي والأنكمومايس ماليري، إحدى الثدييات المُنقرضة المُرتبطة بالليمورية والجنس المُنقرض من الرئيسيات في بداية العصر الجليدي من أواخر الإيوسيني، والقرد القريب، أقرب الفصائل لهم. خلق النقص في أمشاط الأسنان البارزة في السجل الحفري، قبل ما يقرب من 40 فترة مدارية، صراعًا مع دراسات الساعة الجزيئية، التي تعرض اختلاف أقدم بين الليمور وحيوان اللوريس مع وجود شبح لأنساب الرئيسيات الليمورية بأفريقيا.
التراكيب المُتشابهة من حيث التركيب والوظيفة

إن مشط الأسنان، الترتيب الشكلي الخاص للأسنان في الفك السفلي الأمامي، موجود لدى الهباريات الرئيسة غير المُنقرضة، التي تتضمن رئيسيات الليمور وحيوان اللوريس، المعروفة كليًا باسم الليموريات[ملحوظة 1]).[4] هذا البناء التركيبي هو علامة مميزة تساعد على التعرف على ذلك الفرع الحيوي من الرئيسيات.[5][6] توجد السمة الوظيفية في الواكاري الأصلع، وهو نوع من قردة العالم الجديد.[7]
من المُمكن أيضًا أن يتواجد مشط الأسنان في الكولوجو وزبابيات الشجر، حيث إنهما الأقرب إلى الرئيسيات، وذلك على الرغم من اختلاف التراكيب، وهو ما يُعد مثالًا على التطور المُتقارب.[8][9] وبالمثل، فإن الظباء الأفريقية صغيرة أو مُتوسطة الحجم، مثل الإمبالة[10] تمتلك تركيبًا مُشابه، يُشار إليه أحيانًا بجهاز التنظيف السني الجانبي.[11][12] وتمتلك أيضًا حيوانات الوبر الحية والمُنقرضة مشطًا للأسنان، على الرغم من أن عدد الشوكات في المشط يختلف خلال السجل الحفري.[13][14]
منذ حقبة العصر الإيوسيني، أي ما يزيد عن 40 فترة مدارية، امتلكت لقمانيات وكلبيات الشكل الدببية، وهما من أنواع الثدييات المشيمية الأولية، مشطًا للأسنان متطورًا بشكل مستقل.[15][16]
التركيب التشريحي
يتضمن مشط الأسنان لمُعظم الليموريات ستة أسنان تفصل بينهم مسافات، فهم أربعة قواطع وسنتان نابيتان في حالة انبطاح، في حالة ميل إلى الأمام، أمام الفم.[5][17] وللسنة النابية السفلى المُنبطحة نفس شكل القواطع الموجودة بينهم،[18] ولكنهم أكثر قوة وانحناء إلى الأعلى وإلى الداخل، أكثر من القواطع.[15] وفي التسنين الدائم، تظهر الأنياب بعد القواطع.[19] ويكون لتيجان القواطع زاوية باتجاه مائل للأمام،[17] بينما تكون تيجان القواطع والأنياب مُستطيلة ومضغوطة جنبًا إلى جنب.[20] ويكون العرف الرأسي الذي يتبع الأطراف الأمامية لسنة المشط في مُعظم الليموريات على شكل حرف (V)، ويتناقص تدريجيًا من عند خط الوسط.[21] وكنتيجة لإعادة التشكيل السنية هذه، لا تتصل القواطع العليا والسفلى، ودائمًا ما تقل القواطع العليا، أو يتم فقدها بالكامل.[17][22]
قام عالم التشريح، هنري ماري دوكروتي دي بلينفيل، للمرة الأولى بالتعرف على السنتين الجانبيتين من مشط الأسنان الليموري بوصفهم أنياب عام 1840.[23] تستخدم الأسنان النابية عادةً في خرق الأشياء أو مسكها.[24] مع الأسنان النابية السفلية المُعدلة، فإن أول ضرس أمامي يتبع مشط الأسنان يشبه تمامًا السنة نابية الشكل،[5][25] ويقوم بوظائفها.[17] وعادةً ما تكون تلك الضروس الأمامية في حالة التباس مع الأنياب.[26] فعادةً تستقر الأنياب الحقيقية بالفك السفلي أمام الأنياب العليا. بينما في الرئيسيات التي تملك مشطًا للأسنان يقع الضرس الأمامي النابي خلف الأنياب العليا.[17]
تحافظ منطقة تحت اللسان على نظافة مشط الأسنان الليموري، وهي تركيب عضلي مُختص يعمل كفرشاة أسنان لإزالة الشعر والبقايا الأخرى. يمكن لمنطقة تحت اللسان أن تمتد إلى أسفل نهاية اللسان، وتكون مُزودة بنقاط مُسننة ذات قرون تميل على الأسنان الأمامية.[27][28][29]
يختلف تركيب مشط الأسنان بين الليموريات؛[4] فمشط الأسنان لدى الإندرية يكون أقل انبطاحًا،[30] ويتكون من أربع أسنان، بدلًا من ستة. ويمتاز مشط الأسنان لدى الإندرية بأنه أكثر قوة واتساعًا، حيث تكون قواطعه أقصر، بمسافات أكبر بين الأسنان (مسافات بينسنية)، وعرف رأسي أعرض.[21] ليس هناك ما يؤكد ما إذا كان ذلك المشط السني المكون من أربع أسنان يتكون من زوجين من القواطع أو زوج من القواطع وزوج آخر من الأنياب.[5][31] أما عن ليمور الشوكة المُتوج، فيكون مشط أسنانه مضغوطًا أكثر، بمسافات بينسنية أقل. ويمتاز الست أسنان جميعهم بأنهم أكثر طولًا واستقامة، ويشكلوا عرفًا رأسيًا مُتصلًا. بينما في الليموراوات القديمة والسيفاكاوات الأحفورية تم فقد مشط الأسنان، وأكملت القواطع والأنياب تكوينها النموذجي أمام الفم.[32] وقد فقد الآيآي أيضًا مشطه السني، حيث تم استبداله بسنة أمامية مرتفعة متواصلة النمو، شبيهة بقواطع القوارض.[5][33]
يكون لمشط الأسنان لدى الكولوجو تركيب مُختلف تمامًا. فبدلًا من القواطع الفردية والأسنان النابية التي تفصلهم مسافات بسيطة لتعمل كأسنان المشط، يُصبح الطرف القارض للأربع قواطع مُسنن بأكثر عدد ممكن وهو 15 شوكة لكل قاطع.[34] بينما يعمل الناب غالبًا كضرس خلفي.[35] ويتم الحفاظ على نظافة تلك القواطع المُسننة عن طريق الجزء الأمامي من اللسان، والتي تكون مُسننة لتلائم تسنن القواطع.[27] وبالمثل، فإن مشط الأسنان لدى الوبريات يتكون من قواطع مُزودة بالعديد من الشوكات تُسمى المُمشطات. وعلى النقيض من الكولوجو، فحجم الشوكات وشكلها أكثر تماثلًا.[13]
يُشبه مشط الأسنان لدى زبابيات الشجر مشط الأسنان الليموري في أن له مسافات بينسنية لتكوين شوكات المشط. ولكن يحتوي المشط على زوجين فقط من ثلاثة أزواج من القواطع السفلى،[9][34][36] بينما يتم استبعاد الأنياب.[34] وبصفة عامة، فإن القاطعين بمشط الأسنان يمتازان بحجم أكبر.[9] في كلبيات الشكل الدببية المُنقرضة كانت الستة قواطع السفلى كلها جزء من مشط الأسنان.[34] ومشط الأسنان في الظباء الأفريقية يشبه مشط الأسنان في الليموريات بشكل كبير في أنه يتكون من زوجين من القواطع مع زوج من الأنياب.[11]
الوظائف
لمشط الأسنان العديد من الوظائف الحيوية، بوصفه تركيب مُتشابه تركيبيًا في الليموريات، وذلك على الرغم من نمطية شكله ومظهره ظاهريًا.[6] فهو يُستخدم في الأساس كأداة للتزين أو مشط للتنظيف.[37][38] وتستخدم بعض الفصائل مشط الأسنان أيضًا في الحصول على الغذاء[37] أو نزع لحاء الأشجار.[38]
التنظيف

أول من لاحظ الوظيفة الأساسية لمشط الأسنان، وهي التنظيف، كان عالم الطبيعة الفرنسي جورج كوفييه عام 1829. حيث أشار إلى أن الليمور حلقي الذيل يمتلك قواطع سفلى عبارة عن أمشاط حقيقية.[35] وبعد مرور أكثر من مائة عام، ظهرت تساؤلات حول وظيفة التنظيف، حيث كان من الصعب مُلاحظتها. وكان هناك ظن أن المسافات البينسنية صغيرة جدًا بالنسبة للفراء.[39] وقد أظهرت الملاحظات لاحقًا أن الأسنان تُستخدم لهذا الغرض، وأن من المُمكن أن يعلق الشعر بالأسنان بعد عملية التنظيف مُباشرةً، ولكن تتم إزالته لاحقًا بواسطة منطقة تحت اللسان.[40]
في عام 1981، كشف المجهر الفاحص للإلكترونات عن وجود أخاديد أو تصدعات دقيقة في أسنان المشط السني لليموريات. ووُجدت تلك الأخاديد فقط على أطراف الأسنان، في الأسطح المُقعرة بين الأطراف، بالإضافة إلى العرف الخلفي للأسنان. تُشير تلك الأخاديد أن الشعر قد تحرك عدة مرات عبر الأسنان ما بين اتساع 10 و20 ميكرومتر.[ملحوظة 2] وكان بداخل تلك الأخاديد أخاديد أصغر، أقل من 1 ميكرومتر، تكونت بفعل تسلخ الطبقة الجُليدية للشعر.[16]
من بين غير الرئيسيات، تمتاز لقمانيات الشكل المُنقرضة بأنه لديها أخاديد مجهرية في مشطها السني،[16] في حين أن الليمور الفلبيني الطائر لا يمتاز بذلك. يؤدي المشط السني للكولوجو وظيفة مشط الأسنان، ولكن نظرًا لقلة التصدعات بالأسنان، وعدم وجود مُلاحظات لاستخدام مشط الأسنان أثناء عملية التنظيف بالفم، يبدو أن استخدامه مُقتصر على الحصول على الغذاء.[34]
في الظباء الأفريقية، يبدو أن جهاز التنظيف السني الجانبي لا يُستخدم خلال الرعي. فبدلًا من ذلك، تُستخدم خلال التنظيف، عندما تندفع الرأس إلى أعلى في حركة مميزة. فمن الظن أنها تُمشط الفراء وتزيل الطفيليات الخارجية.[11][12]
الشم لدى الليموريات
من المُمكن أيضًا أن يلعب مشط الأسنان لدى الليموريات الرئيسية دورًا ثانويًا في الشم، والذي يمكن أن يتناسب مع النقص في حجم القواطع العليا، التي لم تحظ بدراسة كافية.[41][42] من المُمكن لمشط الأسنان أن يزود الضغط فيعمل على تحفيز الافرازات الغُدية التي تنتشر حينها في الفراء.[43] وعلاوةً على ذلك، فإن النقص في حجم القواطع العليا يُمكن أن يصنع فجوة بين الأسنان، والذي يُعرف بانشقاق الأسنان، التي تصل النثرة، وهي شق في منتصف الأنف الرطبة، بالعضو الميكعي أنفي في سقف الفم. ومن المُمكن أن يسمح ذلك للفيرومونات أن تنتقل بسهولة إلى العضو الميكعي أنفي.[42][44]
الحصول على الغذاء والاستخدامات الأخرى
تستخدم الليمورات الفأرية والسيفاكا والإندرية المشط السني للوصول إلى لب الفاكهة.[16] وتستخدم الليموريات الصغيرة الأخرى مثل ليمور الشوكة المُتوج والليمور القزم مشعر الأذنين والجلاجو، وتحديدًا نوع الجلاجو ومخالب الإبرة، المشط السني لكشط ما تنضحه النباتات مثل المادة الصمغية والنسغ.[16][45] ويستطيع مشط الأسنان لدى ليمور الشوكة المُتوج التكيف بشكل خاص مع تقليل جعل الطعام عالقًا في الأسنان، حيث تقل المسافات البينسنية بشكل كبير.[46] كما يُمكن للكولوجو آكل العشب، الذي يكون من نفس نوع الليمور الفلبيني الطائر، استخدام مشطه السني للحصول على الغذاء.[34]
تستخدم الإندريات مثل السيفاكا مشطها السني لنزع اللحاء أو الخشب الميت، وهو ما يُعرف برفع اللحاء،[16][38][45] وهي العملية التي تقوم بها قبل تمييز الرائحة بواسطة غدة موجودة بالصدر.[47] وهناك ظن بأن التركيب الأقوى من مشطها السني يُساعدها على مُقاومة القوى الضاغطة التي تتعرض لها أثناء عملية رفع اللحاء المُنتظمة.[21]
التطور في الليموريات

كانت أصول مشط الأسنان الليموري، والفرع الحيوي الذي يميزه أساسًا لمُناقشة هامة استمرت لأكثر من قرن. في عام 1920، افترض عالم مستحاثات أسلاف البشر البريطاني ويلفريد كلارك أن مشط الأسنان الذي وُجد في زبابيات الشجر، الذي كان يظن أنهم من الرئيسيات، كان شكلًا مُبكرًا من التركيب السني الذي وُجد في الليموريات. ولأنه قام بدراسة اللوريسيات الحفرية في العصر الميوسيني على أنهم لم يطوروا مشط الأسنان الليموري الحديث بالكامل، فقد ألمح أن الليمور واللوريسيات قد طوروا تلك السمة بطريقة مُستقلة. ولكن تلك الدراسة اختلفت لاحقًا، وأصبحت العلاقة ذات الأصل الواحد بين الليمور واللوريسيات مقبولة الآن.[48]
تُشير الحالة السلفية للتسنين الأمامي بالفك السفلي، التي تقوم على الحفريات الرئيسة للعصر الإيوسيني، أن الرئيسيات السابقة كان ينقصها مشط أسنان مُتباين.[49] كانت مُعظم الهباريات الحفرية في افتقار إلى مشط الأسنان الليموري النمطي. كانت رئيسيات الهباريات السابقة تُعرف جماعيًا باسم أدبيات الشكل .[50] وكانت أدبيات الشكل تُعد مجموعة شبه عرقية، تضم مجموعة كبيرة من أحفاد آخر سلف مُشترك من أفراد المجموعة، وذلك لأن من المُفترض أن الليموريات قد تطورت من إحدى المجموعات العديدة لأدبيات الشكل.[51] وبالنسبة لعلم البيئة، فإن من المُفترض أن تطور مشط الأسنان قد تطلب نظام غذائي آكل للعشب وأوراق الشجر بين أعداد أدبيات الشكل السلفية، حيث إن ذلك من شأنه أن ينتقي للقواطع المُصغرة، التي يُمكن أن تكون بمثابة تكيف مسبق، وهي سمة ذات قيمة تكيفية بديل عما كانت قد اختيرت له، والتي يُمكن أن تُستخدم حينها في التنظيف الشخصي أو الجماعي.[38] وعلى الرغم من ذلك، فإن من الضروري أن احتواء مشط الأسنان على أنياب قد تطلب ظروفا استثنائية، حيث إن الليموريات الكبيرة قد قامت ثانويًا بتغيير القبطواحن النابية لتعويض النقص.[52]
هناك فرضية شائعة حول أصول الفرع الحيوي الليموري، تقول بأنهم قد تطوروا من الأدبيات الأوروبية المعروفة بالأدابيسات.[53] في بعض الأدابيسات تصطف أعراف القواطع السفلى والأنياب لتشكل وحدة نامية فعالة. وقد أشار البليونتولوجي الأمريكي فيليب جنجريتش أن هذا قد ألقى الضوء على تطور مشط الأسنان الليموري.[54] وعلى الرغم من ذلك، لم يتم العثور على أي مشط أسنان ليموري في السجل الحفري للعصر الإيوسيني.[55] كما أن الفكوك السفلية للأدابيسات الأوروبية الموجودة بذلك الوقت لا تُشبه تلك الحالة المُستمدة التي شُوهدت في الليموريات.[56]
هناك ظن حاليًا أن الليموريات قد تطورت في أفريقيا، وأن الهباريات الرئيسية المعروفة باكرًا من أفريقيا هم أعزبيات، من أوائل العصر الإيوسيني،[50] والتي من المُرجح أنها جاءت من الاستعمار الباكر جدًا للكتلة الأرضية العربية الأفريقية في العصر الباليوسيني، من 66 إلى 55 فترة مدارية.[57] وُجدت سلالة الليموريات التي تشمل الليمور الجبلي والأنكمومايس ماليري في أفريقيا، وتُؤرخ من 50 إلى 48 فترة مدارية، وكانت تختلف كثيرًا عن أدبيات الشكل الأوروبية.[50] ولكن على الرغم من ذلك، كانوا يفتقرون إلى مشط الأسنان.[57][58] تُشير هذه السلالة من الليموريات إلى وجود أصل مُشترك مُبكر مع السيركامونيات من خارج أوروبا.[59] [ملحوظة 3] وارتكازًا على وجود أسنان سفلية منبطحة، كان هناك ظن أن القرد القريب، وهي رئيسة حفرية وُجدت في رواسب أواخر عصر الإيوسين في مُنخفض الفيوم بمصر، لها صلة وثيقة بالليموريات.[60][61] يُعتبر كل من القرد القريب والأنكمومايس ماليري والقرد القريب من أصناف شقيقة لليموريات الرئيسة.[57]
التضارب في التأريخ
على الرغم من أن الليموريات الأصلية، مثل الليمور الجبلي، من المُحتمل أن تكون مُتعاصرة مع رئيسيات ذات مشط سني مُرتبطة بها من حوالي 50 إلى 48 فترة مدارية، فإن السجل الحفري الأفريقي المُتناثر يُشير إلى أن التنوع في مشط الأسنان تم من حوالي 52 إلى 40 فترة مدارية، وذلك وفقًا لعالم البليوأنثروبولوجيا الفرنسي مارك جودينوت.[50] وهذا من شأنه أن يخلق صراعًا مع تقديرات الساعة الجزيئية التي توصل إليها عالم الأنثروبولوجيا التطوري آن يودر وآخرون، التي تُتنبأ بالاختلاف الليموري اللوريسي المُؤرخ بين 61 و90.8 فترة مدارية.[62]
في عام 2001، قاوم اكتشاف الليمورات البكتية، وهي رئيسية حفرية من باكستان يرجع تاريخها إلى العصر الأوليغوسيني واعتُقد في البداية أنها من الليموريات فأرية الشكل، التاريخ النشوئي لليموريات.[63] وعلى الرغم من ذلك، فقد تبين لاحقًا أنها نوع من أدبيات الشكل الرئيسة، وليست ليمورًا.[64]
تضاعف تقريبًا الحد الأدنى للتقدير البليونتولوجي لتشعب الليمورات واللوريسيات عندما ظهرت اكتشافات إضافية من الجلاجو الأصلي، وهي الغلاغويات الصحراوية، واللوريسيات الأصلية أو التاجية، وهي القارونيسيات، ترجع من 37 إلى 40 فترة مدارية على التوالي، في شمالي مصر خلال فترة 2000.[65][66] والقارونيسيات هي أقدم رئيسية حفرية يظهر لديها مشط أسنان ليموري مميز.[67] ويُوحي ذلك، بالإضافة إلى أدبيات الشكل الإفريقية الأخرى مثل الأنكمومايس ماليري، بوجود شبح أنساب أقدم لليموريات بأفريقيا.[57][68]
الوظيفة الأصلية لمشط الأسنان لدى الليموريات
كان الضغط الانتقائي الذي شكل مشط الأسنان الليموري الأصلي موضوعًا لمُناقشات هامة من فترة 1970. ويُمكن القول بأن الدليل يدعم وظيفة التنظيف أو الحصول على الغذاء أو الاثنين معًا.[69] في بدايات فترة 1900، كان هناك جدل بنسبة أقل، حيث كان يُرى أن وظيفة التنظيف هي الوظيفة الأساسية، حيث كانت الرئيسيات في حالة افتقار إلى المخالب اللازمة لتمشيط الفراء على نحو كافٍ، وذلك على الرغم من أن السعالي البدائية الرئيسة، مثل الهباريات والأبخص، تمتلك على الأقل مخلب تنظيف واحد على كل قدم للتعويض.[35] بشكل عام، يُعد التنظيف، في هيئة تمشيط الفراء، الوظيفة الأساسية والدور الأصلي لمشط الأسنان الليموري. ولكن التغييرات اللاحقة في المورفولوجيا عبر العديد من سلاسل النسب قد قام بتبديل وظيفتها وإخفاء وظيفتها الأصلية.[70]
كانت الفرضية التي تقول أن مشط الأسنان قد تطور من أجل وظيفة الحصول على الغذاء ترتكز على ملاحظات لأصنوفة ليمورية حديثة، مثل الليمورات فأرية الشكل، وبالأخص ليمورات الشوكة المُتوجة والليمور القزم مشعر الأذنين والجلاجو، التي تُوضح كشط الأسنان لنضاحة النباتات. هذا بالإضافة إلى السيفاكا التي تقوم بعملية نزع اللحاء. كان كل ما سبق يُعد من الأشكال الأولية بين الهباريات الحية، وهو بدوره ما يُشير إلى الليموريات الأولى أظهرت سلوكًا مُشابهًا.[45] وأيضًا تم التركيز على الضغط الانتقائي القوي من بيئة التغذية، الموجودة في التسنين الأمامي، بالاعتماد على تسنين أمامي علوي مُتخصص يُمكن رؤيته في ليمور الكوالا المُنقرض حديثًا، ميجالادبيس. إذا كان لبيئة التغذية هذا التأثير العميق على شكل التسنين الأمامي، فإن من المُمكن للتطور المُتقارب أن يُوضح التشابهات المرئية بين القواطع السفلى المضغوطة لمشط الأسنان الليموري وتكيف التغذية الإفرازي في جنس المارموست.[71]
على النقيض، أكدت فرضية التنظيف أن جميع الليموريات تستخدم مشطها السني للتنظيف، والأسنان الطويلة النحيفة تكاد لا تتناسب مع الضغط الآلي الناتج عن نزع اللحاء واستخراج الغذاء.[71] كما أن المسافات البينسنية الموجودة في مُعظم الليموريات تدعم تمشيط الفراء، وهي تعزز أيضًا من نمو البكتيريا وتسوس الأسنان إذا استُخدمت في إطعام التغذية. ومما يؤيد ذلك، وجود المسافة البينسنية المصغرة في إطعام التغذية لليموريات.[72] وعلاوةً على ذلك، يوفر الناب الموجود في مشط الأسنان مسافة بينسنية إضافية لتمشيط الفراء.[73] وحتى سلوك الليموريات الصغيرة يشير إلى أن التنظيف يلعب دورًا أكثر أهمية في استخدام مشط الأسنان من الحصول على الغذاء.[74]
مُلاحظات
- على الرغم من أن العلاقة ذات الأصل الواحد بين الليمورات واللوريسيات مقبولة بشكل كبير، فإن اسم فرعهما الحيوي ليس كذلك. فكلمة هباريات قد تم استخدامها كضمادة سلة مهملات لرئيسيات النسناسيات غير بسيطة الأنف، التي لا يُظهر مُعظمها مشط الأسنان المُميز. وكانت الليموريات تُسبب مشكلة لكونها عضو من الرئيسيات الحية والحفرية، التي تضمنت ليمورات مدغشقر والأدبيات الأوروبية، بينما تم استبعاد اللوريسيات منها. تُستخدم كلمة ليموريات الشكل هنا لأنها تنشأ من تصنيف واحد شائع يجمع الرئيسيات التي تمتلك مشطًا للأسنان تحت رتبة واحدة، تجمع الأدبيات المُنقرشة التي لا تمتلك مشطًا للأسنان تحت رتبة أخرى، وكليهما تحت رتبة الهباريات.
- أشار عالم الأنثروبولوجيا الطبيعية الأمريكي جيفري إتش شوارتز أن حيوان الأبخص يمتلك مشطًا سنيًا على درجة عالية من الاشتقاق، ومُتشابه تركيبيًا مع مشط الأسنان الليموري، وقد أشار إلى ذلك استنادًا إلى أنماط التآكل الدقيقة السنية التي هي إحدى خصائص التمشيط السني، بالإضافة إلى التشابهات بين القاطع الأمامي المُتماوج أفقيًا، وشكل السنة النابية الجانبية في مشط الأسنان الليموري. ولكن أوضحت الدراسات حول تسنين حيوان الأبخص أن بعض فصائل الأبخص فقط هي من يظهر لديها التحزيز المُميز على القواطع، في حين أن جميع الفصائل التي تم فحصها ذُكر أنها تستخدم أسنانها الأمامية أثناء التنظيف، مُشيرًا إلى أن وجود التحزيزات أو غيابها يُمكن أن يتأثر بتكرار التنظيف أو سُمك الفراء أو المُباعدة البينسنية. تم توثيق التمشيط السني، كوظيفة للأسنان بوجود مشط الأسنان أو عدم وجوده، في الرئيسيات المُبكرة في أواخر العصر الإيوسيني. كان الليمور الميت، وهو نوع أوروبي وأقرب الفصائل للأبخص، في حاجة إلى مشط أسنان، ولكنها أظهرت علامات من التآكل الدقيق المنسوب إلى التمشيط السني.
- كانت الرئيسيات الأحفورية فرعًا مبكرًا من رئيسيات الهباريات التي شغلت نص الكرة الشمالي في أوائل فترة الباليوجين. نشأت الرئيسيات الأحفورية في آسيا، وكانت تندرج تحت فصيلة الدبيات النوثية، وهي رئيسيات مُنقرضة من أمريكا الشمالية، تشبه الليمورات الحديثة.
مصادر
- Toothcomb نسخة محفوظة 23 سبتمبر 2018 على موقع واي باك مشين.
- Lemurs: Ecology and Adaptation نسخة محفوظة 24 سبتمبر 2018 على موقع واي باك مشين.
- The Prosimians: John G. Fleagle, in Primate Adaptation and Evolution (Third Edition), 2013 نسخة محفوظة 17 ديسمبر 2019 على موقع واي باك مشين.
- Cuozzo & Yamashita 2006, p. 68.
- Cuozzo & Yamashita 2006, p. 73.
- Szalay & Seligsohn 1977, p. 76.
- Eaglen 1980, pp. 278 & 280.
- Wood Jones 1918, pp. 345–346.
- Eaglen 1980, pp. 279–280.
- Mooring, McKenzie & Hart 1996, p. 970.
- McKenzie 1990, p. 117.
- Mooring, McKenzie & Hart 1996, p. 966.
- Stevens et al. 2009, p. 974.
- Function of the mandibular tooth comb in living and extinct mammals نسخة محفوظة 17 ديسمبر 2019 على موقع واي باك مشين.
- Eaglen 1980, p. 277.
- Rose, Walker & Jacobs 1981, p. 583.
- Ankel-Simons 2007, p. 235.
- Ankel-Simons 2007, p. 235
- Eaglen 1980, p. 281.
- Osman Hill 1953, pp. 52–53.
- Szalay & Seligsohn 1977, p. 78.
- Homologies of the toothcomb نسخة محفوظة 24 سبتمبر 2018 على موقع واي باك مشين.
- Eaglen 1980, p. 279.
- Ankel-Simons 2007, p. 234.
- Osman Hill 1953, p. 53.
- Wood Jones 1918, p. 345.
- Wood Jones 1918, p. 347.
- Osman Hill 1953, p. 73.
- Ankel-Simons 2007, pp. 421–423.
- Ankel-Simons 2007, p. 254.
- Ankel-Simons 2007, p. 233.
- Szalay & Seligsohn 1977, pp. 78–79.
- Ankel-Simons 2007, p. 257.
- Rose, Walker & Jacobs 1981, p. 585.
- Wood Jones 1918, p. 346.
- Wood Jones 1918, p. 346. Sargis 2004, p. 58.
- Ankel-Simons 2007, p. 238.
- Rosenberger 2010, p. 291.
- Roberts 1941, p. 236.
- Roberts 1941, pp. 236–237.
- Cuozzo & Yamashita 2006, p. 72.
- Cuozzo & Yamashita 2006, pp. 74–76.
- Rosenberger & Strasser 1985, p. 73.
- Asher 1998, p. 355.
- Szalay & Seligsohn 1977, pp. 76–77.
- Szalay & Seligsohn 1977, p. 80.
- Mittermeier et al. 2010, p. 572.
- Rosenberger & Strasser 1985, p. 74.
- Godinot 2006, p. 455.
- Godinot 2006, p. 446.
- Godinot 2006, pp. 446–447.
- Szalay & Seligsohn 1977, p. 77.
- Tattersall 2006, p. 9.
- Godinot 2006, p. 447.
- Rasmussen & Nekaris 1998, pp. 256–258.
- Godinot 2006, pp. 455–456.
- Seiffert et al. 2005, p. 11400.
- Godinot 2006, p. 452.
- Godinot 2006, pp. 453–455.
- Simons & Rasmussen 1994, p. 9946.
- Rasmussen & Nekaris 1998, p. 261.
- Godinot 2006, p. 458.
- Tattersall 2006, pp. 8–9.
- Godinot 2006, pp. 447–449.
- Jaeger et al. 2010, p. 1095.
- Seiffert et al. 2005, p. 11396.
- Tattersall 2006, p. 7.
- Seiffert, Simons & Attia 2003, pp. 423–424.
- Cuozzo & Yamashita 2006, pp. 73–74.
- Szalay & Seligsohn 1977, p. 75.
- Rosenberger & Strasser 1985, p. 75.
- Szalay & Seligsohn 1977, pp. 80–81.
- Cuozzo & Yamashita 2006, p. 74.
- Eaglen 1980, p. 276.
مراجع
- Ankel-Simons، F. (2007). Primate Anatomy (ط. 3rd). Academic Press. ISBN:978-0-12-372576-9.
{{استشهاد بكتاب}}: الوسيط|ref=harvغير صالح (مساعدة) - Asher، R. J. (1998). "Morphological diversity of anatomical strepsirrhinism and the evolution of the lemuriform toothcomb". American Journal of Physical Anthropology. ج. 105 ع. 3: 355–367. DOI:10.1002/(SICI)1096-8644(199803)105:3<355::AID-AJPA5>3.0.CO;2-Q. PMID:9545077.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Beard، C. (2004). The Hunt for the Dawn Monkey: Unearthing the Origins of Monkeys, Apes, and Humans. University of California Press. ISBN:978-0-520-23369-0. مؤرشف من الأصل في 2020-11-25.
{{استشهاد بكتاب}}: الوسيط|ref=harvغير صالح (مساعدة) - Cartmill، M. (2010). "Chapter 2: Primate Classification and Diversity". في Platt، M.؛ Ghazanfar، A (المحررون). Primate Neuroethology. Oxford University Press. ص. 10–30. ISBN:978-0-19-532659-8.
{{استشهاد بكتاب}}: الوسيط|ref=harvغير صالح (مساعدة) - Cuozzo، F. P.؛ Yamashita، N. (2006). "Chapter 4: Impact of Ecology on the Teeth of Extant Lemurs: A Review of Dental Adaptations, Function, and Life History". في Gould، L.؛ Sauther، M. L. (المحررون). Lemurs: Ecology and Adaptation. Springer. ص. 67–96. ISBN:978-0-387-34585-7. مؤرشف من الأصل في 2022-04-07.
{{استشهاد بكتاب}}: الوسيط|ref=harvغير صالح (مساعدة) - Eaglen، R. H. (1980). "Toothcomb homology and toothcomb function in extant strepsirhines". International Journal of Primatology. ج. 1 ع. 3: 275–286. DOI:10.1007/BF02692275.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Godinot، M. (2006). "Lemuriform origins as viewed from the fossil record". Folia Primatologica. ج. 77 ع. 6: 446–464. DOI:10.1159/000095391. PMID:17053330.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Jaeger، J. J.؛ Beard، K. C.؛ Chaimanee، Y.؛ Salem، M.؛ Benammi، M.؛ Hlal، O.؛ Coster، P.؛ Bilal، A. A.؛ Duringer، P.؛ Schuster، M.؛ Valentin، X.؛ Marandat، B.؛ Marivaux، L.؛ Métais، E.؛ Hammuda، O.؛ Brunet، M. (2010). "Late middle Eocene epoch of Libya yields earliest known radiation of African anthropoids" (PDF). Nature. ج. 467 ع. 7319: 1095–1098. DOI:10.1038/nature09425. PMID:20981098. مؤرشف من الأصل (PDF) في 2013-03-08.
- McKenzie، A. A. (1990). "The ruminant dental grooming apparatus". Zoological Journal of the Linnean Society. ج. 99 ع. 2: 117–128. DOI:10.1111/j.1096-3642.1990.tb00564.x.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - قالب:LoM3
- Mooring، M.؛ McKenzie، A. A.؛ Hart، B. L. (1996). "Grooming in impala: Role of oral grooming in removal of ticks and effects of ticks in increasing grooming rate" (PDF). Physiology & Behavior. ج. 59 ع. 4–5: 965–971. DOI:10.1016/0031-9384(95)02186-8.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة)[وصلة مكسورة] - Musser، G. G.؛ Dagosto، M. (1987). "The identity of Tarsius pumilus, a pygmy species endemic to the montane mossy forests of central Sulawesi" (PDF). American Museum Novitates. ج. 2867: 1–53. ISSN:0003-0082. مؤرشف من الأصل (PDF) في 2014-07-27.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Osman Hill، W. C. (1953). Primates Comparative Anatomy and Taxonomy I—Strepsirhini. Edinburgh Univ Pubs Science & Maths, No 3. Edinburgh University Press. OCLC:500576914.
{{استشهاد بكتاب}}: الوسيط|ref=harvغير صالح (مساعدة) - Rasmussen، D. T.؛ Nekaris، K. A. I. (1998). "Evolutionary history of lorisiform primates" (PDF). Folia Primatologica. ج. 69 ع. Suppl. 1: 250–285. DOI:10.1159/000052716. مؤرشف من الأصل (PDF) في 2020-11-25.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Roberts، D. (1941). "The dental comb of lemurs". Journal of Anatomy. ج. 75 ع. Pt 2: 236–238. PMC:1252661. PMID:17104854.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Rose، K. D.؛ Walker، A.؛ Jacobs، L. L. (1981). "Function of the mandibular tooth comb in living and extinct mammals". Nature. ج. 289 ع. 5798: 583–585. DOI:10.1038/289583a0. PMID:7007889.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Rosenberger، A. L. (2010). "Chapter 14: Adaptive profile versus adaptive specialization: Fossils and gummivory in early primate evolution". في Burrows، A. M.؛ Nash، L. T (المحررون). The Evolution of Exudativory in Primates. Springer. ص. 273–295. ISBN:978-1-4419-6660-5. مؤرشف من الأصل في 2020-07-21.
{{استشهاد بكتاب}}: الوسيط|ref=harvغير صالح (مساعدة) - Rosenberger، A. L.؛ Strasser، E. (1985). "Toothcomb origins: Support for the grooming hypothesis". Primates. ج. 26: 73–84. DOI:10.1007/BF02389048.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Sargis، E. J. (2004). "New views on tree shrews: The role of Tupaiids in primate supraordinal relationships" (PDF). Evolutionary Anthropology: Issues, News, and Reviews. ج. 13 ع. 2: 56–66. DOI:10.1002/evan.10131. مؤرشف من الأصل (PDF) في 2021-02-12.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Schmid، P. (1983). "Front Dentition of the Omomyiformes (Primates)". Folia Primatologica. ج. 40: 1–10. DOI:10.1159/000156088.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Schwartz، J. H. (2003). "Chapter 3: How close are the similarities between Tarsius and other primates?". في Wright، P. C.؛ Simons، E. L.؛ Gursky، S (المحررون). Tarsiers: Past, Present, and Future. Rutgers University Press. ص. 50–96. ISBN:978-0-8135-3236-3.
{{استشهاد بكتاب}}: الوسيط|ref=harvغير صالح (مساعدة) - Seiffert، E. R.؛ Simons، E. L.؛ Attia، Y. (2003). "Fossil evidence for an ancient divergence of lorises and galagos". Nature. ج. 422 ع. 6930: 421–424. DOI:10.1038/nature01489. PMID:12660781.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Seiffert، E. R.؛ Simons، E.L.؛ Ryan، T. M.؛ Attia، Y. (2005). "Additional remains of Wadilemur elegans, a primitive stem galagid from the late Eocene of Egypt" (PDF). Proceedings of the National Academy of Sciences. ج. 102 ع. 32: 11396–11401. DOI:10.1073/pnas.0505310102. PMC:1183603. PMID:16087891. مؤرشف من الأصل (PDF) في 2015-09-24.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Simons، E. L.؛ Rasmussen، D.T. (1994). "A remarkable cranium of Plesiopithecus teras (Primates, Prosimii) from the Eocene of Egypt" (PDF). Proceedings of the National Academy of Sciences. ج. 91 ع. 21: 9946–9950. DOI:10.1073/pnas.91.21.9946. PMC:44934. مؤرشف من الأصل (PDF) في 2015-09-24.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Stevens، N. J.؛ O'Connor، P. M.؛ Roberts، E. M.؛ Gottfried، M. D. (2009). "A hyracoid from the Late Oligocene Red Sandstone Group of Tanzania, Rukwalorax jinokitana (gen. and sp. nov.)" (PDF). Journal of Vertebrate Paleontology. ج. 29 ع. 3: 972. DOI:10.1671/039.029.0302. مؤرشف من الأصل (PDF) في 2015-12-10.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Szalay، F. S.؛ Seligsohn، D. (1977). "Why did the strepsirhine tooth comb evolve?". Folia Primatologica. ج. 27: 75–82. DOI:10.1159/000155778.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة) - Tattersall، I. (2006). "Chapter 1: Origin of the Malagasy strepsirhine primates". في Gould، L.؛ Sauther، M.L (المحررون). Lemurs: Ecology and Adaptation. Springer. ص. 3–20. ISBN:978-0-387-34585-7. مؤرشف من الأصل في 2022-04-07.
{{استشهاد بكتاب}}: الوسيط|ref=harvغير صالح (مساعدة) - Wood Jones، F. (1918). "The sublingua and the plica fimbriata". Journal of Anatomy. ج. 52 ع. 4: 345–353. PMC:1262842. PMID:17103846.
{{استشهاد بدورية محكمة}}: الوسيط|ref=harvغير صالح (مساعدة)
وصلات خارجية
- مشط أسنان الليامر.
- أصول ووظائف مشط الأسنان عند الليامر.
- دور مشط الأسنان عند الثدييات المنقرضة والباقية.
 بوابة رئيسيات
بوابة رئيسيات بوابة تشريح
بوابة تشريح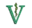 بوابة بيطرة
بوابة بيطرة بوابة علم الحيوان
بوابة علم الحيوان بوابة ثدييات
بوابة ثدييات