قالب قراءة مفتوح
إطار القراءة المفتوح أو قالب القراءة المفتوح (بالإنجليزية: open reading frame) هو جزء من إطار قراءة يملك قابلية أن يُترجم إلى ناتج جيني (بروتين أو رنا غير مشفر وظيفي)، وهو مفهوم في علم الأحياء الحاسوبي يُعرَّف على أنه امتداد مستمر من الكودونات (الرامزات) يبدأ بكودون بداية (عادة AUG) وينتهي بكودون توقف (عادة UAA أو UAG أو UGA) ويُستخدم كوسيلة للتنبؤ الأولي بالجينات.[1] جاءت كلمة مفتوح من ترجمة كلمة Open في العبارة "open for translation" أي له قابلية للترجمة وهي إشارة إلى احتمال وجود جين مشفر للبروتين. [2]
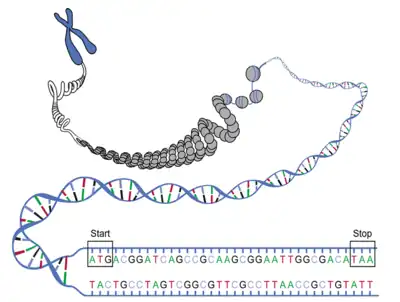
يوجد اختلاف في بنية الجين بين بدائيات وحقيقيات النوى، فجينات حقيقيات النوى تحتوي على عدة إكسونات وتتم إزالة الإنترونات أثناء التوصيل للحصول على رنا رسول ناضج. وفي سياق البحث عن الجينات، لا ينطبق تعريف إطار القراءة المفتوح المحدد بكودونات البداية والنهاية سوى على جينات بدائيات النوى البسيطة أو جزيئات الرنا الرسول الناضجة وليس على الدنا الجينومي لحقيقيات النوى، لأن الإنترونات قد تحتوي على كودونات توقف أو تسبب انزياحات بين إطارات القراءة، لذلك ينص تعريف حديث شامل على أن إطار القراءة المفتوح هو تسلسل نوكليوتيدات قابل للقسمة على ثلاثة يحده كودونا توقف.[2][3] وهذا التعريف الشامل مفيد في سياق النسخوميات والميتاجينوميات، حيث قد لا يتواجد كودون توقف و/أو كودون بداية في التسلسل المتحصل عليه، ومثل إطار القراءة المفتوح ذلك يمكن أن يعود إلى أجزاء من جين بدل الجين بأكمله.
يحد إطارات القراءة المفتوحة من الجانبين مناطق غير مشفرة هي: المنطقة 5' غير المترجمة والمنطقة 3' غير المترجمة، وهي مناطق من الجين تُنسخ في الرنا الرسول أثناء النسخ لكنها لا تُترجم إلى تسلسل أحماض أمينية أثناء الترجمة. وتحتوي هذه المناطق على عناصر تنظيمية مهمة لترجمة الجين مثل المحفزات والمحسنات والكاتمات.
غالبا ما تحتوي جزيئات الرنا الرسول البكتيري على عدة إطارات قراءة في نسخة واحدة، وفي مثل هذه الحالة يسمى ذلك الجين بالمشغل ويسمى الرنا الرسول الخاص به عديد السيسترون.
الأهمية البيولوجية
أحد الاستخدامات الشائعة لإطارات القراءة المفتوحة هي كونها أحد الأدلة المساعدة في التنبؤ بالجينات. غالبا ما تُستعمل إطارات القراءة المفتوحة الطويلة - إلى جانب أدلة أخرى- في التحديد الأولي للمناطق المشفرة للبروتين أو المناطق المشفرة للرنا الوظيفي في تسلسل الدنا. تواجد إطار قراءة مفتوح في منطقة ما لا يعني دائما أن تلك المنطقة مشفرة للبروتين. على سبيل المثال: في تسلسل دنا عشوائي له نسبة متساوية من كل نوكليوتيد، سيتواجد كودون توقف كل 21 كودون.[4] قد تبحث خوارزمية بسيطة للتنبؤ بالجينات الخاصة ببدائيات النوى على كودون بداية يتبعه إطار قراءة مفتوح يكون طويلا كفاية ليشفر بروتينا نموذجيا، يكون فيه استخدام الكودونات في تلك المنطقة مطابقا لتواتر استخدام الكودونات الخاص بذلك الكائن في المناطق المشفرة للبروتين.[4] وعليه، يقول بعض الكتّاب أن إطار القراءة المفتوح يجب أن يكون له طول أدنى قدره 100 كودون[5] أو 150 كودون.[4] وحتى إطار القراءة المفتوح الطويل بحد ذاته ليس دليلا حاسما على وجود جين.[4] من جهة أخرى، تم إثبات أن بعض إطارت القراءة المفتوحة القصيرة (sORFs) التي تعوز السمات الكلاسيكية الخاصة بالجينات المشفرة للبروتين (في كل من جزيئات الرنا غير المشفر والرنا الرسول) يمكن أن تشفر ببتيدات وظيفية.[6] من المعروف أن 50% من المناطق 5' غير المترجمة في جزيئات الرنا الرسول الخاصة بالثدييات تحتوي على إطارات قراءة مفتوحة قصيرة.[7] وحوالي 64-75% من مواقع بدء ترجمة إطارات القراءة المفتوحة القصيرة المحددة تجريبيا هي مواقع محفوظة في جينومات البشر والفئران ويمكن أن يدل ذلك على أن هذه العناصر لها وظيفة.[8] مع ذلك، لا تتواجد إطارت القراءة المقتوحة القصيرة عادة سوى في أنواع قليلة من جزيئات الرنا الرسول وتتجنب الاصطفاء، يمكن أن يكون الانحفاظ الكبير لمواقع البدء مرتبطا بأماكنها داخل محفزات الجينات الخاصة بها. توجد حالة مثل هذه في جين SLAMF1.[9]
اقرأ أيضا
المراجع
- "Open reading frame". U.S. National Library of Medicine. 19 أكتوبر 2015. مؤرشف من الأصل في 2016-01-28. اطلع عليه بتاريخ 2015-10-22.
- Sieber P, Platzer M, Schuster S (2018). "The Definition of Open Reading Frame Revisited". Trends Genet. ج. 34 ع. 3: 167–170. DOI:10.1016/j.tig.2017.12.009. PMID:29366605. مؤرشف من الأصل في 2020-11-01.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: أسماء متعددة: قائمة المؤلفين (link) - Claverie, J.-M. (1997) Computational methods for the identification of genes in vertebrate genomic sequences. Hum. Mol. Genet. 6, 1735–1744.
- Deonier، Richard؛ Simon Tavaré؛ Michael Waterman (2005). Computational Genome Analysis: an introduction. شبغنكا. ص. 25. ISBN:978-0-387-98785-9.
- Claverie, J.-M., Poirot, O., Lopez, F. (1997) The difficulty of identifying genes in anonymous vertebrate sequences. Comput. Chem. 21 203-214
- Zanet، J.؛ Benrabah، E.؛ Li، T.؛ Pelissier-Monier، A.؛ Chanut-Delalande، H.؛ Ronsin، B.؛ Bellen، H. J.؛ Payre، F.؛ Plaza، S. (2015). "Pri sORF peptides induce selective proteasome-mediated protein processing". Science. ج. 349 ع. 6254: 1356–1358. DOI:10.1126/science.aac5677. ISSN:0036-8075. PMID:26383956. S2CID:206639549.
- Wethmar، Klaus؛ Barbosa-Silva، Adriano؛ Andrade-Navarro، Miguel A.؛ Leutz، Achim (1 يناير 2014). "uORFdb—a comprehensive literature database on eukaryotic uORF biology". Nucleic Acids Research. ج. 42 ع. D1: D60–D67. DOI:10.1093/nar/gkt952. ISSN:0305-1048. PMC:3964959. PMID:24163100. مؤرشف من الأصل في 2017-10-18.
- Lee, Sooncheol; Liu, Botao; Lee, Soohyun; Huang, Sheng-Xiong; Shen, Ben; Qian, Shu-Bing (11 Sep 2012). "Global mapping of translation initiation sites in mammalian cells at single-nucleotide resolution". Proceedings of the National Academy of Sciences (بالإنجليزية). 109 (37): E2424–E2432. DOI:10.1073/pnas.1207846109. ISSN:0027-8424. PMC:3443142. PMID:22927429.
- Schwartz، Anton M.؛ Putlyaeva، Lidia V.؛ Covich، Milica؛ Klepikova، Anna V.؛ Akulich، Kseniya A.؛ Vorontsov، Ilya E.؛ Korneev، Kirill V.؛ Dmitriev، Sergey E.؛ Polanovsky، Oleg L. (1 أكتوبر 2016). "Early B-cell factor 1 (EBF1) is critical for transcriptional control of SLAMF1 gene in human B cells". Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms. ج. 1859 ع. 10: 1259–1268. DOI:10.1016/j.bbagrm.2016.07.004. PMID:27424222.
- Rolf Knippers: Molekulare Genetik. 8. neubearbeitete Auflage. Georg Thieme Verlag, Stuttgart u. a. 2001, ISBN 3-13-477008-3.
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي