غشاء خلوي
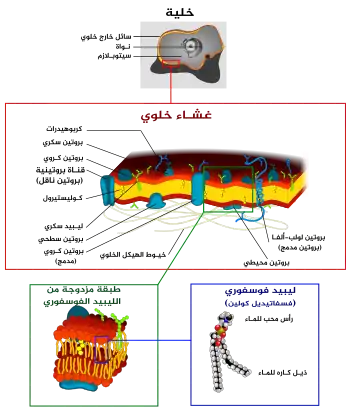
الغشاء الخلوي[2] أو الغشاء البلازمي هو غشاء شفاف يُحيط بالخلية ويفصلها عن البيئة الخارجية .[3] يتكون الغشاء البلازمي من ليبيد ثنائي الطبقة ذو نفاذية اختيارية .[4] تترتب الليبيدات بشكل فسيفسائي. تلعب مكونات الغشاء الخلوي دورًا هامًا في مجموعة واسعة من العمليات الخلوية. في نفس الوقت يمكن أن يعمل كنقطة اتصال بين الهيكل الخلوي والجدار الخلوي كما في الخلايا النباتية. ينظم الغشاء الخلوي دخول وخروج الجزيئات من وإلى الخلية ، بالاضافة إلى احتواءِه على مستقبلات لمجموعة واسعة من الهرمونات والإشارات الخلوية. [5][6][7]
| غشاء خلوي | |
|---|---|
| الاسم العلمي Cell membrane | |
 | |
 غشاء خلوي | |
| تفاصيل | |
| نوع من | غشاء [1] |
| معرفات | |
| ترمينولوجيا هستولوجيكا | H1.00.01.0.00011، وH1.00.01.1.00001 |
| FMA | 63841 |
| ن.ف.م.ط. | A11.284.149 |
| ن.ف.م.ط. | D002462 |
تاريخ
أدى اكتشاف روبرت هوك للخلايا عام 1665 إلى اقتراح نظرية الخلية ، دحض هوك النظرية القائلة بأن جميع الخلايا تحتوي على جدار خلوي صلب. أمكن ملاحظة الخلايا النباتية فقط في ذلك الوقت. ركز علماء الميكروسكوب على جدار الخلية لأكثر من 150 عامًا حتى تم إحراز تقدم في الفحص المجهري. في أوائل القرن التاسع عشر ، تم التعرف على الخلايا على أنها كيانات منفصلة وغير متصلة ومربوطة بجدران خلوية فردية بعد أن تم اكتشاف إمكانية فصل الخلايا النباتية. امتدت هذه النظرية لتشمل الخلايا الحيوانية. بحلول النصف الثاني من القرن التاسع عشر ، لم يكن الفحص المجهري متقدمًا بما يكفي للتمييز بين أغشية الخلايا وجدران الخلايا. ومع ذلك ، حدد بعض علماء الميكروسكوب بشكل صحيح في هذا الوقت أنه في حين أنه غير مرئي ، يمكن استنتاج أن أغشية الخلايا كانت موجودة في الخلايا الحيوانية بسبب الحركة داخل الخلايا للمكونات داخليًا ولكن ليس خارجيًا وأن الأغشية لم تكن مكافئة لجدار الخلية نباتية. كما تم الاستدلال على أن أغشية الخلايا ليست مكونات حيوية لجميع الخلايا. دحض الكثيرون وجود غشاء خلوي لا يزال حتى نهاية القرن التاسع عشر. في عام 1890 ، ذكر تحديث لنظرية الخلية أن أغشية الخلايا كانت موجودة ، لكنها مجرد هياكل ثانوية. لم يتم التعرف على أغشية الخلايا إلا بعد دراسات لاحقة حول التناضح والنفاذية. في عام 1895 ، اقترح إرنست أوفرتون أن أغشية الخلايا مصنوعة من الدهون.
خلقت فرضية الطبقة الدهنية ثنائية الطبقة ، التي اقترحها جورتر وجريندل في عام 1925 ، تكهنات في وصف بنية طبقة غشاء الخلية الثنائية استنادًا إلى الدراسات البلورية وملاحظات فقاعات الصابون. في محاولة لقبول أو رفض الفرضية ، قام الباحثون بقياس سمك الغشاء حين استخرج هؤلاء الباحثون الدهون من خلايا الدم الحمراء البشرية وقاسوا مساحة السطح التي سيغطيها الدهن عند انتشاره على سطح الماء. خلايا الدم الحمراء في الثدييات تفتقر إلى كل من النوى والعضيات السيتوبلازمية ، وغشاء البلازما هو الهيكل الوحيد المحتوي على الدهون في الخلية. وبالتالي ، يمكن افتراض أن جميع الدهون المستخرجة من الخلايا قد استقرت في أغشية البلازما في الخلايا. تم حساب نسبة مساحة سطح الماء المغطاة بالدهن المستخلص إلى مساحة السطح المحسوبة لخلايا الدم الحمراء تمثل 2: 1 (تقريبًا) وخلصوا إلى أن غشاء البلازما يحتوي على طبقة دهنية ثنائية.
في عام 1925 ، حدد فريك أن سمك أغشية خلايا الدم الحمراء والخميرة يتراوح بين 3.3 و 4 نانومتر . تم التشكيك في اختيار ثابت العزل الكهربائي المستخدم في هذه الدراسات ولكن الاختبارات المستقبلية لم تستطع دحض نتائج التجربة الأولية. بعد إختراع المجهر الضوئي تم قياس الأغشية الرقيقة جدًا من خلال مقارنة شدة الضوء المنعكس بكثافة معيار الغشاء ذي السماكة المعروفة. يمكن للأداة حل السماكات التي تعتمد على قياسات الأس الهيدروجيني ووجود بروتينات الغشاء التي تراوحت من 8.6 إلى 23.2 نانومتر ، مع القياسات السفلية التي تدعم فرضية طبقة ثنائية الدهون. في وقت لاحق من الثلاثينيات ، تم تطوير نموذج هيكل الغشاء بالاتفاق العام ليكوننموذج جزيئي ضئيل من دافسون ودانيلي ( 1935). اعتمد هذا النموذج على دراسات التوتر السطحي بين الزيوت وبيض شوكيات الجلد . نظرًا لأن قيم التوتر السطحي بدت أقل بكثير مما كان متوقعًا بين الزيت والماء ، فقد افتُرِض أن بعض المواد كانت مسؤولة عن خفض التوترات السطحية في سطح الخلايا. تم اقتراح أن طبقة ثنائية الدهون تقع بين طبقتين من البروتين الرقيق. أصبح نموذج الجزيئات الصغيرة شائعًا على الفور وسيطر على دراسات غشاء الخلية على مدار الثلاثين عامًا التالية ، حتى أصبح ينافس نموذج الفسيفساء السائل لسينجر ونيكلسون.
على الرغم من النماذج العديدة لغشاء الخلية التي تم اقتراحها قبل النموذج الفسيفساء ، إلا أنه تظل النموذج الأساسي لغشاء الخلية بعد فترة طويلة من إنشائها في السبعينيات. على الرغم من تحديث نموذج الفسيفساء السائل إلا أن الأساسيات ظلت ثابتة "الغشاء عبارة عن طبقة ثنائية من الدهون تتكون من رؤوس خارجية محبة للماء وداخلية كاره للماء" حيث يمكن للبروتينات أن تتفاعل مع الرؤوس المحبة للماء من خلال التفاعلات القطبية ، ولكن البروتينات التي تمتد على الطبقة الثنائية تحتوي كليًا أو جزئيًا على أحماض أمينية كارهة للماء تتفاعل مع الجزء الداخلي للدهون غير القطبية. نموذج الفسيفساء السائل لم تقدم فقط تمثيلًا دقيقًا لميكانيكية الأغشية ، بل عززت أيضًا دراسة القوى الكارهة للماء ، والتي ستتطور لاحقًا إلى تحديد وصفي للجزيئات البيولوجية الكبيرة .
لما يقرب من قرنين من الزمان ، شوهدت الأغشية ولكن تم تجاهلها . لم يكن حتى في القرن العشرين أهمية للغشاء الخلوي حتى تم الاعتراف بها. أخيرًا ، اكتشف العالمان جورتر وجريندل (1925) أن الغشاء مكون بشكل رئيسي من الدهون. عُززت فكرة أن الهيكل التركيبي يجب أن يكون في تشكيل يحاكي الطبقات. تم دراسة هذه الفرضية بشكل أكبر إلا أن تم العثور عليها من خلال مقارنة مجموع أسطح الخلايا وأسطح الدهون ، تم تقدير نسبة 2: 1 ؛ وبالتالي ، توفير الأساس الأول للبنية ثنائية الطبقة المعروفة اليوم.
التركيب
تحتوي أغشية الخلايا على مجموعة متنوعة من الجزيئات البيولوجية ، لا سيما الدهون والبروتينات. يتغير الغشاء الخلوي باستمرار ويغير من سيولته تبعًا للتغيرات البيئية حي. يتغير خلال المراحل المختلفة من تطور الخلية.تتغير كمية الكوليسترول في غشاء الخلية العصبية الأولية للإنسان ، ويؤثر هذا التغيير في التركيب على السيولة خلال مراحل التطور. على الرغم من أن تركيز مكونات الغشاء في المرحلة المائية منخفض لذا تكون مكونات الغشاء الخلوي مستقرة ولها قابلية منخفضة للذوبان في الماء ، هناك تبادل للجزيئات بين المرحلتين الدهنية والمائية.
الدهون
يتكون غشاء الخلية من ثلاث فئات من الدهون الدهون الفوسفورية ، والشحميات السكرية ، والستيرولات . تعتمد كمية كل منها على نوع الخلية ، ولكن في معظم الحالات تكون الفسفوليبيدات هي الأكثر وفرة ، وغالبًا ما تساهم بأكثر من 50٪ من إجمالي الدهون في أغشية البلازما. تمثل الجليكوليبيدات فقط كمية قليلة تبلغ حوالي 2٪ بينما تشكل الستيرولات النسبة المتبقية. في دراسة على خلايا الدم الحمراء ، وجد أن 30٪ من غشاء البلازما عبارة عن دهون. ومع ذلك ، بالنسبة لغالبية الخلايا حقيقية النواة ، يكون تكوين أغشية البلازما حوالي نصف دهون ونصف بروتينات.
تحتوي السلاسل الدهنية في الفسفوليبيدات والجليكوليبيدات عادة على عدد زوجي من ذرات الكربون ، عادةً ما بين 16 و 20 ذرة كربون. الأحماض الدهنية ذات الـ 16، 18 ذرة كربون هي الأكثر شيوعًا. قد تكون الأحماض الدهنية مشبعة أو غير مشبعة .درجة عدم التشبع في سلاسل الأحماض الدهنية لها تأثير كبير على سيولة الغشاء حيث أن الدهون غير المشبعة تخلق التواء ، مما يمنع الأحماض الدهنية من التماسك معًا بإحكام ، وبالتالي تقليل درجة حرارة الانصهار ومنه تزيد من سيولة العشاء الخلوي. قدرة بعض الكائنات الحية على تنظيم سيولة أغشية خلاياها عن طريق تغيير تكوين الدهون يسمى التكيف المثلي .
يتم تثبيت الغشاء بأكمله عبر تفاعل غير تساهمي للذيول الكارهة للماء ،ولكن الهيكل مائع تمامًا وغير ثابت في مكانه. في ظل الظروف الفسيولوجية ، تكون جزيئات الفسفوليبيد في غشاء الخلية في الحالة البلورية السائلة . وهذا يعني أن جزيئات الدهون تظهر انتشارًا جانبيًا سريعًا على طول الطبقة التي توجد فيها. ومع ذلك ، تعتبر أطواف الدهون والكافولايات أمثلة على التركيبات الغنية بالكوليسترول في غشاء الخلية. يوجد جزء من الدهون في اتصال مباشر مع البروتينات الغشائية المتكاملة ، والتي ترتبط بإحكام بسطح بروتين يسمى الغلاف الدهني الحلقي ؛ يتصرف كجزء من مركب البروتين.
الكوليسترول
يوجد الكوليسترول في الخلايا الحيوانية عادةً منتشرًا بدرجات متفاوتة في العشاء الخلوي ، خصوصًا بين الفراغات غير المنتظمة الموجودة بين ذيول الدهون الكارهة للماء ، حيث يضفي مرونة للغشاء الخلوي. تختلف كمية الكوليسترول في الأغشية البيولوجية بين الكائنات الحية وأنواع الخلايا وحتى في خلايا النوع الواحد. ينظم الكوليسترول ، وهو مكون رئيسي في أغشية البلازما الحيوانية سيولة الغشاء الكلي ، مما يعني أن الكوليسترول يتحكم في مقدار حركة مكونات غشاء الخلية المختلفة بناءً على تركيزاته.في درجات الحرارة المرتفعة ، يمنع الكوليسترول حركة سلاسل الأحماض الدهنية الفسفورية ، مما يؤدي إلى تقليل نفاذية الجزيئات الصغيرة وتقليل سيولة الغشاء. والعكس صحيح بالنسبة لدور الكوليسترول في درجات الحرارة المنخفضة. يتم تنظيم كمية الكوليسترول على أغشية الخلايا، وبالتالي تركيز ، استجابة لدرجة الحرارة . في درجات الحرارة الباردة ، يتدخل الكوليسترول في تفاعلات سلسلة الأحماض الدهنية. يعمل الكوليسترول كمضاد للتجمد ، ويحافظ على سيولة الغشاء. وهذا يفسر سبب وجد الكوليسترول بكثرة في الحيوانات التي تعيش في البيئات الباردة والمتجمدة أكثر منها في الحيوانات التي تعيش في المناطق الدافئة. أغشية الخلايا النباتية تفتقر إلى الكوليسترول ، وتحوي أغشيتها عوضًا عنه مادة شبيهه تسمى ستيرول لها نفس وظيفة الكوليسترول في الخلايا الحيوانية.
الفسفوليبيدات في الحويصلات الدهنية
الحويصلات الدهنية أو الجسيمات الشحمية عبارة عن جيوب كروية تقريبًا محاطة بطبقة ثنائية من الدهون. تُستخدم هذه الهياكل في المختبرات لدراسة تأثيرات المواد الكيميائية في الخلايا عن طريق إيصال هذه المواد الكيميائية مباشرة إلى الخلية ، بالإضافة إلى الحصول على مزيد من التبصر في نفاذية غشاء الخلية. تتشكل الحويصلات الدهنية والجسيمات الشحمية عن طريق تعليق مادة دهنية أولاً في محلول مائي ثم تحريك الخليط من خلال الصوتنة ، مما يؤدي إلى تكوين حويصلة. عن طريق قياس معدل التدفق من داخل الحويصلة إلى المحلول المحيط ، يسمح هذا للباحثين بفهم نفاذية الغشاء بشكل أفضل. يمكن أن تتكون الحويصلات من جزيئات وأيونات داخل الحويصلة عن طريق تكوين الحويصلة بالجزيء أو الأيون المرغوب الموجود في المحلول. يمكن أيضًا دمج البروتينات في الغشاء من خلال إذابة البروتينات المرغوبة في وجود المنظفات وربطها بالفوسفوليبيدات التي يتكون فيها الجسيم الشحمي. هذه الخاصية تمكن الباحثين من فحص وظائف بروتينات الأغشية المختلفة.
الكربوهيدرات
تتمثل الكربوهيدرات في الغشاء البلازمي ، بالبروتينات السكرية ،والجليكوليبيدات. الكربوهيدرات مهمة في عملية التعرف والتواصل بين الخلايا. توجد الكربوهيدرات على سطح الخلية حيث تتعرف على الخلايا المضيفة وتتشارك المعلومات ، كما يمكن لبعض الفيروسات استخدام هذه الكربوهيدرات كمستقبلات لها على أسطح الخلايا المستهدفة. بالنسبة للجزء الأكبر ، لا يحدث ارتباط بالجليكوزيل على الأغشية داخل الخلية ؛ يحدث الارتباط بالجليكوزيل بشكل عام على السطح خارج الخلية لغشاء البلازما.يعتبر الجلايكوكاليكس(glycocalyx) جزيء مهم في جميع الخلايا ، خصوصًا الخلايا الظهارية المبطنة للمرات الهضمية. تشير البيانات الحديثة إلى أن للجلايكوكاليكس القدرة على المشاركة في التصاق الخلية ، كما يلعب دورًا في توجيه الخلايا الليمفاوية إلى مكان العدوى.
البروتينات
يحتوي غشاء الخلية على نسبة كبيرة من البروتينات ، حوالي 50٪ من حجم الغشاء . هذه البروتينات مهمة للخلية لأنها مسؤولة عن الأنشطة البيولوجية المختلفة. تتكون بروتينات الغشاء من ثلاثة أنواع رئيسية: البروتينات المتكاملة ، والبروتينات المحيطية ، والبروتينات الدهنية.
البروتينات المتكاملة هي بروتينات أمفيباثية. تتضمن أمثلة البروتينات المتكاملة القنوات الأيونية ومضخات البروتون والمستقبلات المقترنة ببروتين g. تسمح القنوات الأيونية للأيونات غير العضوية مثل الصوديوم أو البوتاسيوم أو الكالسيوم أو الكلور بالانتشار عكس تدرجها الكهروكيميائي. يتم التحكم في السلوك الكهربائي للخلايا (أي الخلايا العصبية) بواسطة القنوات الأيونية. مضخات البروتون هي مضخات بروتينية مدمجة في طبقة ثنائية الدهون تسمح للبروتونات بالمرور عبر الغشاء عن طريق الانتقال من سلسلة جانبية من الأحماض الأمينية إلى أخرى. تستخدم العديد من العمليات الحيوية مضخات البروتون مثل نقل الإلكترونات وتوليد ATP . المستقبلات المقترن بالبروتين G عبارة عن سلسلة أحادية متعددة الببتيد متموجه الشكل (سبع موجات) على سطح الغشاء الخلوي، تستجيب لجزيئات والإشارة (أي الهرمونات والناقلات العصبية). تُستخدم المستقبلات المقترنة بالبروتين G في عمليات مثل إرسال الإشارات من خلية إلى أخرى ، وتنظيم إنتاج cAMP ، وتنظيم القنوات الأيونية.
يعتبر غشاء الخلية موقعًا مهمًا للتواصل بين الخلية والخلايا الأخرى.وعلى هذا النحو توجد مجموعة كبيرة ومتنوعة من المستقبلات البروتينية التي تستقبل الإشارات من الخلايا المجاورة لتعمل معًا وفق تناسق دقيق . يمكن أن تشمل وظائف البروتينات الغشائية أيضًا التلاصق بين الخلية والخلية المجاورة، والتعرف على السطح ، والتلاصق مع الهيكل الخلوي الداخلي ، والنشاط الأنزيمي ، أو نقل المواد عبر الغشاء.
الوظيفة
يقوم الغشاء الخلوي أيضا بإحاطة الهيولى وفصلها فيزيائيا عن بقية المكونات الخارج خلوية، وبهذا يقوم بمهمة جدار فاصل. هذا الحاجز قادر على تنظيم خارج وداخل الخلية الحية باعتباره نصف نفوذ أو نفوذ نوعيا، وهو غشاء مُنفِذ يعمل عمل غشاء نصف ناضح وفي نفس الوقت له القابلية الاختيارية للمواد الداخلة والخارجة من خلاله، كما تساهم في حالات كثيرة جزيئات ناقلة موجودة ضمن الغشاء البلازمي في عملية نقل المواد عبر الغشاء وتتم هذه العملية بصرف كميات كبيرة من الطاقة الكيميائية. وبالنظر لكون الغشاء البلازمي رقيق جدًا؛ لذلك لا يمكن تمييزه بسهولة باستخدام المجهر الضوئي ويلاحظ في بعض الخلايا أن الغشاء البلازمي محاط بطبقات واقية أكثر سمكًا بحيث يمكن تمييزها بالمجهر فمثلًا معظم الخلايا النباتية تمتلك جدارًا سيليلوزيًا سميكًا يغطي ويدعم الغشاء البلازمي والذي يطلق عليه اسم الجدار الخلوي Cell Wall أما خلايا الحيوانات فبعضها محاط بمواد قوية. إن للجدار الخلوي الوظائف المهمة للخلية غير أنه لا يلعب أي دور في عملية عبور المواد وانتقالها من وإلى الخلية.
النقل عن طريق تكوين الحويصلات
لأغشية بعض الخلايا القدرة على إحاطة بعض المواد وتكوين حويصلات غشائية حيث عن طريقها يتم ادخال وإخراج هذه المواد من وإلى الخلية.
- أولا: الإدخال الخلوي Endocytosis: يتم أخذ الأجسام إلى داخل الخلية عن طريق غشاء البلازما بعدة بطرق منها:
- الالتهام الخلوي (أو البلعمة) Phagocytosis يمثل الالتهام الخلوي هضم الأجسام الصلبة من الخلية بواسطة الفعالية الطبيعية لغشاء البلازما. هذه الظاهرة يمكن ملاحظتها في الأميبا حيث تعمل على مد أقدام كاذبة حول الدقائق المطلوب هضمها ثم تحتوي هذه الدقائق في داخل الخلية وتتكون فجوة كبيرة نسبيًا تنطلق إلى داخل الخلية وأن عمل بعض خلايا الدم البيضاء يكون مشابهًا لعمل الأميبا والتي تساعد الجسم في الوقوف ضد المواد الغريبة حيث أن كريات الدم البيضاء Leucocytes لها القابلية لهضم البكتريا بواسطة الأكياس الملتهمة وكذلك فضلات الخلية وأجسام كبيرة أخرى.
- الشرب الخلوي Pinocytosis: يمثل الشرب الخلوي احتواء المواد السائلة إلى داخل الخلية بطريقة تشبه البلعمة وقد يدخل البروتين بهذه الطريقة أيضًا حيث تمتز المواد Adsorbed عند سطح البلازما ثم يحدث لف داخلي Infolding للغشاء ناتجًا في تكوين كيس يحتوي على الدقائق المطلوب هضمها وبعدها فأن هذه المواد تتحرر بطريقة ما من الكيس إلى داخل الخلية وأن الغشاء الذي كان محيطًا للدقيقة قد يصبح جزءًا من الشبكة الإندوبلازمية ويمكن تلخيص العملية وذلك بتكوين أصابع غشائية خارجية تنحني بعدها إلى الخلف لتلتحم في النهاية بغشاء الخلية ومرة ثانية مكونة بذلك فجوة غذائية تحتوي بداخلها على المادة الغذائية.
- اللقف الخلوي Rhopheocytosis: وهذه الآلية في الإدخال الخلوي خاصة لنقل كمية كبيرة من المواد مثل السيتوبلازم مع محتوياته من خلية إلى خلية أخرى حيث تتضمن العملية تكوين فجوات في سطح الخلية دون وجود تقديرات سابقة في السطح حيث تظهر الخلية في هذه العملية كأنها تشفط المواد المحيطة بها كالشرب الخلوي.
- ثانيا: الإخراج الخلوي Exocytosis
ويمكن تقسيم هذه العملية إلى عدة اقسام هي:
أ- الإفراز الكلي Holocrine Secretion : ويتضمن هذا الإفراز ملئ الخلية بالناتج الإفرازي ثم تحرر الخلية برمتها كجسم إفرازي وبعدها تضمحل الخلية محررة محتوياتها وتمثل الغدد الدهنية لجلد اللبائن انموذجًا لهذا النوع من الإفراز.
ب- الإفراز الجزئي Eccrine Secretion : وهو عكس الإفراز الكلي إذ تبدأ العملية ببناء البروتينات السكرية بواسطة الشبكة الإندوبلازمية الخشنة المحببة) Granular Endoplasmic Reticulum ثم ترزم بهيئة أجسام محددة بأغشية ثم تحرر محتوياتها داخل تجويف بواسطة التحام الغشاء الموجود حول الجسم بالغشاء الخلوي وتنتج عن هذه العملية انخفاضات مؤقتة تنشأ عند سطح الخلية وفي حالة الإفراز الشديد ينشأ خيط من فجوات مرتبطة مع بعضها البعض وبواسطة هذه الوسائل ينبذ الإفراز إلى الخارج ومن الأمثلة لهذا النوع كثير من الغدد ذات الإفراز الخارجي والداخلي كالبنكرياس والخلايا الأمامية للغدة النخامية والخلايا الدرقية. وأن التحام غشاء الجسم الإفرازي بالغشاء البلازمي يؤدي إلى أن يصبح غشاء الجسم الإفرازي جزءًا من الغشاء البلازمي.
3- الإفراز القمي Apocrine Secretion : إن هذا الإفراز يلاحظ في الغدد تحت الفكية للأرنب Submandibular sweet gland ولقد لوحظت عمليات مختلفة من الإفراز حيث يندفع التجويف السطحي للخلية إلى الخارج ليكون بروزات ثانوية وأشكال كروية متصلة بالخلية بواسطة سويق رفيع بعدها تكون طبقة كثيفة من السيتوبلازم على عرض الساق تفصل تدريجيًا الجسم المخزون ويصبح طليقًا في التجويف ويمكن مشاهدة هذه العملية أيضًا في الغدد اللبنية حيث يفرز الدهن بواسطة آلية الإفراز القمي.
4- الإفراز الثنائي Diacrine Secretion : في هذا النوع من الإفراز تتكون أجسام إفرازية محاطة بأغشية كما في الإفراز الجزئي ولكن بدلًا من تحررها بالتحامها بالغشاء البلازمي فإن الناتج الإفرازي إما ينتشر إولًا عبر غشاء الجسم الإفرازي ثم عبر الغشاء البلازمي أو تنتشر أجزاء غشاء الجسم الإفرازي والإفراز المتحرر عبر السيتوبلازم القمي وغشاء البلازما، إن هذه العملية لا تتضمن زيادة أو نقصان في محيط سطح الخلية كما أنها نادرة الحدوث نسبيًا.
- ثالثا: الانتشار الحر Free Diffusion
تتحرك الكثير من المواد من خلال الأغشية بأسلوب الانتشار الحر، كما تشير الدراسات حيث تتناسب نسبة هذا الانتشار طرديًا مع نسبة ذوبان تلك المواد في الليبيد. يستثنى الماء من هذه القاعدة؛ وذلك لأن جزيئاته تنتشر بحرية خلال الغشاء بانتظام وسرعة، حيث اقترح بأن الأغشية تحتوي على ثقوب (8-10) انكستروم تبطن بجزيئات محبة للماء حيث تكون هذه الفتحات ذات سعة كافية لدخول جزيئات الماء في حين تلاقي جزيئات أخرى صعوبة للدخول من خلالها.
المراجع
- Gene Ontology release 2019-10-07 (ط. 2019-10-07)، 7 أكتوبر 2019، QID:Q70472376
- نزار مصطفى الملاح، معجم الملاح في مصطلحات علم الحشرات (بالعربية والإنجليزية)، الموصل: جامعة الموصل، ص. 162، QID:Q118929029
- html Kimball's Biology pages, Cell Membranes نسخة محفوظة 03 سبتمبر 2014 على موقع واي باك مشين. [وصلة مكسورة]
- Alberts B, Johnson A, Lewis J؛ وآخرون (2002). Molecular Biology of the Cell (ط. 4th). New York: Garland Science. ISBN:0-8153-3218-1. مؤرشف من الأصل في 2009-02-14.
{{استشهاد بكتاب}}: Explicit use of et al. in:|مؤلف=(مساعدة)صيانة الاستشهاد: أسماء متعددة: قائمة المؤلفين (link) - Budin، Itay؛ Devaraj، Neal K. (29 ديسمبر 2011). "Membrane Assembly Driven by a Biomimetic Coupling Reaction". Journal of the American Chemical Society. 134 (2): 751–753. DOI:10.1021/ja2076873. مؤرشف من الأصل في 2015-05-31. اطلع عليه بتاريخ 2012-02-18.
- Staff (25 يناير 2012). "Chemists Synthesize Artificial Cell Membrane". علم يوميا. مؤرشف من الأصل في 2017-10-20. اطلع عليه بتاريخ 2012-02-18.
- Staff (26 يناير 2012). "Chemists create artificial cell membrane". ريموند كرزويل. مؤرشف من الأصل في 2017-10-20. اطلع عليه بتاريخ 2012-02-18.
ضبط استنادي | |
|---|---|
| دولية | |
| وطنية | |
| أخرى | |
 بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي