عقد قاعدية
العقد القاعدية (بالإنجليزية: Basal Ganglia) هي مجموعة من النوى تحت القشرية، مختلفة المنشأ، الموجودة في دماغ الفقاريات. توجد العديد من الاختلافات لدى الإنسان وغيره من الرئيسيات، بشكل رئيسي في انقسام الكرة الشاحبة إلى منقطة داخلية وأخرى خارجية، وفي انقسام الجسم المخطط. تتمركز النوى القاعدية في قاعدة الدماغ الأمامي وقمة الدماغ المتوسط. تتميز العقد القاعدية باتصالات وثيقة مع القشرة المخية، والمهاد وجذع الدماغ، بالإضافة إلى عدد من مناطق الدماغ الأخرى. ترتبط العقد القاعدية بمجموعة من الوظائف المتنوعة، بما في ذلك التحكم في الحركات الإرادية، والتعلم الإجرائي، والتعلم الاعتيادي، والتعلم الإشراطي، وحركات العين، والمعرفة والشعور.[1][2]
| عقد قاعدية | |
|---|---|
| الاسم العلمي nuclei basales | |
 | |
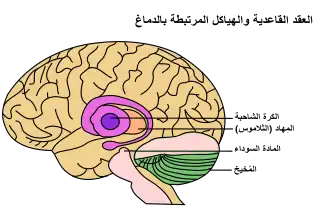 عقد قاعدية مؤشرة في أعلى اليمين عقد قاعدية مؤشرة في أعلى اليمين | |
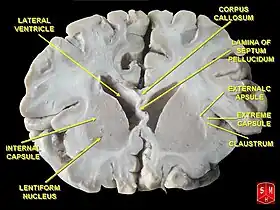 عقد قاعدية في الفص الجبهي من الدماغ عقد قاعدية في الفص الجبهي من الدماغ | |
| تفاصيل | |
| اختصار | BrainInfoType |
| الاختصار | BrainInfoType |
| نوع من | كيان تشريحي معين |
| جزء من | المخ |
| ترمينولوجيا أناتوميكا | 14.1.09.501 |
| FMA | 84013 |
| معلومات عصبية | braininfo |
| UBERON ID | 0010011، و0002420 |
| نيوروليكس | Basal ganglia |
| ن.ف.م.ط. | A08.186.211.730.885.105 |
| ن.ف.م.ط. | D001479 |
| دورلاند/إلزيفير | 12580456 |
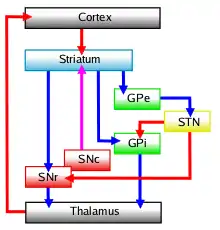
تتألف النوى القاعدية بشكل رئيسي -كما هو محدد وظيفيًا- من الجسم المخطط، الذي يتكون من الجسم المخطط الظهري (النواة الذنبية والبطانة) والجسم المخطط البطني (النواة المتكئة والحديبة الشمية)، والكرة الشاحبة، والشاحبة البطنية، والمادة السوداء ونواة أسفل المهاد. يتميز كل من هذه المكونات بتنظيم عصبي كيميائي وتشريحي داخلي معقد. يتلقى المكون الأكبر، الجسم المخطط (الظهري والبطني)، المدخلات الواردة من مناطق الدماغ المختلفة التي تتجاوز العقد القاعدية، بالإضافة إلى أنه يرسل المخرجات إلى المكونات الأخرى للعقد القاعدية. تتلقى الكرة الشاحبة المدخلات من الجسم المخطط، وترسل المخرجات المثبطة إلى عدد من المناطق المرتبطة بالحركة. تُعد المادة السوداء مصدر المدخلات المخططية للناقل العصبي الدوبامين، الذي يلعب دورًا هامًا في وظيفة العقد القاعدية. تتلقى نواة أسفل المهاد المدخلات من الجسم المخطط والقشرة المخية بشكل رئيسي وترسل إلى الكرة الشاحبة.[3]
تشير النظريات الشائعة إلى دور العقد القاعدية الأساسي في اختيار الفعل – أي المساعدة في اختيار إحدى السلوكيات المحتملة العديدة لتنفيذها في وقت ما. بعبارة أكثر دقة، تتمثل الوظيفة الأساسية للعقد القاعدية في التحكم في نشاطات المناطق القشرية الحركية وقبل الحركية وتنظيمها بهدف أداء الحركات الإرادية بسلاسة. تظهر الدراسات التجريبية امتلاك العقد القاعدية تأثيرًا مثبطًا على عدد من الأنظمة الحركية، إذ يؤدي تحرير هذا التثبيط إلى تعطيل نشاط النظام الحركي. يتأثر «التحويل السلوكي» الذي يحدث في العقد القاعدية بالإشارات الواردة من مختلف أجزاء الدماغ، بما في ذلك قشرة فص الجبهة، التي تلعب دورًا جوهريًا في الوظائف التنفيذية. تشير إحدى الفرضيات أيضًا إلى أن دور النوى القاعدية غير محصور في اختيار الفعل الحركي، بل يشمل أيضًا اختيار الأفعال المعرفية. تجسد النماذج الحاسوبية لاختيار الفعل في العقد القاعدية هذه الفرضية.[4][5][6]
تمتلك العقد القاعدية دورًا هامًا للغاية في وظيفة الدماغ الطبيعي وسلوكه. يؤدي الخلل الوظيفي في هذه العقد إلى مجموعة واسعة من الحالات العصبية بما في ذلك اضطرابات الحركة والتحكم السلوكي، بالإضافة إلى العجز المعرفي المشابه للعجز الناجم عن تلف قشرة فص الجبهة. تشمل الاضطرابات السلوكية كلًا من متلازمة توريت، والاضطراب الوسواسي القهري والإدمان.[7] تشمل اضطرابات الحركة الأكثر شيوعًا مرض باركنسون؛ الذي ينطوي على تدهور الخلايا المنتجة للدوبامين في المادة السوداء؛ ومرض هنتنغتون، الذي يضم بشكل رئيسي تلف الجسم المخطط؛ وخلل التوتر وبشكل أقل شيوعًا الحركية الرقصية النصفية. تمتلك العقد القاعدية قسمًا نطاقيًا مؤلفًا من عدد من المكونات ذات الأسماء البارزة: النواة المتكئة، والشاحبة البطنية والمنطقة السقيفية البطنية (في تي إيه). يشير عدد معتبر من الدلائل إلى لعب القسم النطاقي دورًا مركزيًا في تعلم المكافأة بالإضافة إلى المعرفة ووظيفة الفص الجبهي، من خلال المسار الوسطي الطرفي من «في تي إيه» إلى النواة المتكئة التي تستخدم الناقل العصبي الدوبانين والمسار الوسطي القشري. يُعتقد أن آلية عمل عدد من المخدرات المسببة للإدمان، بما في ذلك الكوكايين والأمفيتامين والنيكوتين، متمثلة في رفع فعالية إشارات الدوبامين. تشير الأدلة أيضًا إلى دور فرط نشاط الإسقاط الدوباميني للمنطقة السقيفية البطنية في الفصام.[8]
الوظيفة
حركات العين
يمثل دور العقد القاعدية في التحكم في حركات العين واحدة من وظائف هذه العقد المدروسة بشكل مكثف. تتأثر حركة العين بشبكة واسعة من المناطق الدماغية التي تتلاقى في منطقة من الدماغ المتوسط معروفة باسم الأكيمة العلوية (إس سي). تمثل الأكيمة العلوية بنية عديدة الطبقات، إذ تشكل طبقاتها خرائط شبكية ثنائية الأبعاد للفضاء البصري. تقود «دفعة» من النشاط العصبي في الطبقات العميقة للأكيمة العلوية حركات العين الموجهة نحو نقطة مطابقة في الفضاء.[9]
تتلقى الأكيمة العلوية إسقاطات مثبطة قوية من العقد القاعدية، التي تنشأ في الجزء الشبكي للمادة السوداء (إس إن آر). غالبًا ما تطلق العصبونات الموجودة في الجزء الشبكي للمادة السوداء معدلات مرتفعة بشكل مستمر، لكنها تصبح «معلقة» عند بدء حركة العين، ما يحرر الأكيمة العلوية من التثبيط. ترتبط جميع حركات العين بجميع أنواعها مع «التعليق» في الجزء الشبكي للمادة السوداء؛ مع ذلك، قد ترتبط عصبونات الجزء الشبكي المفردة بشكل وثيق وقوي أكثر مع بعض حركات العين مقارنة بغيرها من العصبونات. تظهر العصبونات في بعض أجزاء النواة الذنبية أيضًا نشاطًا مرتبطًا بحركات العين. نظرًا إلى إطلاق الغالبية العظمى من الخلايا الذنبية في معدلات منخفضة، يظهر هذا النشاط بشكل دائم تقريبًا كزيادة في معدل الإطلاق. نتيجة لذلك، تبدأ حركات العين بتنشيط النواة الذنبية، التي تثبط الجزء الشبكي للمادة السوداء بواسطة الإسقاطات غابية الفعل المباشرة، التي تزيل بدورها تثبيط الأكيمة العلوية.
دورها في التحفيز
أظهر الدوبامين خارج الخلوي في العقد القاعدية ارتباطًا مع الحالة التحفيزية لدى القوارض، إذ ارتبطت المستويات العالية مع حالة الاكتفاء، والمستويات المتوسطة مع السعي والمستويات المنخفضة مع النفور. تتأثر دوائر العقد القاعدية النطاقية كثيرًا بالدوبامين خارج الخلوي. يؤدي ازياد مستوى الدوبامين إلى حدوث تثبيط في الشاحبة البطنية، والنواة السويقية الداخلية والجزء الشبكي للمادة السوداء، ما يسبب زوال التثبيط في المهاد. يفسر هذا النموذج المؤلف من المسارين المباشر «دي 1» وغير المباشر «دي 2» عدم فعالية الناهضات الانتقائية لكل مستقبل في حدوث المكافأة، إذ يتطلب زوال التثبيط نشاط المسارين على حد سواء. يؤدي زوال تثبيط المهاد إلى تنشيط قشرة فص الجبهة والجسم المخطط البطني، الذي يُعد انتقائيًا لزيادة نشاط «دي 1» ما ينتهي بالمكافأة. تشير الأدلة من دراسات الفيزيولوجيا الكهربائية لدى الإنسان وغيره من الرئيسيات إلى دور البنى الأخرى للعقد القاعدية، مثل الكرة الشاحبة الداخلية ونواة أسفل المهاد، في معالجة المكافأة.[10][11]
اتخاذ القرار
اقتُرح نموذجان اثنان حول آلية عمل العقد القاعدية، إذ يتمثل النموذج الأول في توليد العقد القاعدية للأفعال بواسطة «ناقد» متوضع في الجسم المخطط البطني إلى جانب تقديرها للقيمة، مع تنفيذ هذه الأفعال بواسطة «ممثل» متوضع في الجسم المخطط الظهري. يتمثل النموذج الثاني في عمل العقد القاعدية كآلية انتقائية، إذ تتولد الأفعال في القشرة المخية ثم يقع الاختيار عليها بالاستناد إلى السياق بواسطة العقد القاعدية. تلعب حلقة «سي بي جي تي سي» دورًا في خفض المكافأة، إذ يزداد الإطلاق في حالة المكافأة غير المتوقعة أو الأكبر مقارنة بالمكافأة المتوقعة. دعمت إحدى المراجعات الفكرة القائلة أن القشرة المخية لها دور في أفعال التعلم بغض النظر عن نتائجها، بينما تمتلك العقد القاعدية دورًا في اختيار الأفعال الملائمة بالاستناد إلى تعلم الخطأ والتجربة القائمة على المكافأة المرتبطة.[12]
الذاكرة العاملة
اقتُرح أن العقد القاعدية مسؤولة عن تحديد ما يُسمح له بدخول الذاكرة العاملة. تشير إحدى الفرضيات إلى دور المسار المباشر (الانطلاق أو الاستثارة) في السماح بدخول المعلومات إلى «بّي إف سي»، حيث تبقى مستقلة عن المسار، مع ذلك، تقترح نظرية أخرى استمرار المسار المباشر في الارتداد من أجل بقاء المعلومات في «بّي إف سي». اقتُرح أن المسار القصير غير المباشر، الذي يعمل في آلية دفع وسحب ضد المسار المباشر، مسؤول عن إغلاق البوابة إلى «بّي إف سي».[13]
أهمية سريرية
مرض العقد القاعدية هو مجموعة من اضطرابات الحركة التي تنتج عن إما الإفراط في الإنتاج من العقد القاعدية إلى المهاد أو ناتجة عن عدم كفاية الإنتاج. تنشأ الاضطرابات الحركية الناجمة عن إفراز مفرط من العقد القاعدية، والذي يثبط الناتج من المهاد إلى القشرة، وبالتالي يحد من الحركة الطوعية. تنتج اضطرابات فرط الحركة عن انخفاض الإنتاج من العقد القاعدية إلى المهاد، مما لا يؤدي إلى تثبيط كافي للتوقعات المهادية للقشرة وبالتالي يعطي حركات غير مضبوطة / غير إرادية.[14]
القائمة التالية تحتوي على أمراض لها علاقة بالعقد القاعدية:
- إدمان
- كنع (حركات موجية مستمرة بالأطراف)
- اضطراب نقص الانتباه مع فرط النشاط (ADHD)
- تشنج الجفن
- صرير الأسنان
- شلل دماغي
- رقاص (مرض)
- خلل التوتر
- متلازمة فار
- متلازمة اللكنة الأجنبية (FAS)
- داء هنتنغتون
- يرقان نووي
- متلازمة لش-نيهان
- اضطراب اكتئابي [15]
- اضطراب وسواسي قهري[16][17]
- اضطراب القلق [17]
- باندز
- مرض باركنسون
- خلل النطق التشنجي
- تأتأة[18]
- رقاص سيدنهام
- خلل الحركة المتأخر
- متلازمة توريت
- داء ويلسون
التركيب

يتم تصنيف الجهاز العصبي المركزي البشري في كثير من الأحيان على أساس الحويصلات البدائية الثلاث الأصلية التي يتطور منها: تتكون هذه الحويصلات الأولية في التطور الطبيعي للأنبوب العصبي للجنين وتشمل في البداية الدماغ الأمامي، الدماغ المتوسط والدماغ الخلفي. في اتجاه (من الرأس إلى الذيل).[19] وفي وقت لاحق من تطوير الجهاز العصبي يتحول كل جزء منهم إلى مكونات أصغر. خلال التطور، يتم توجيه الخلايا لتشكيل العقد القاعدية من قبل الانحرافات العقدية الأفقية والإنسية. يوضح الجدول التالي هذا التصنيف التطوري ويتبعه الهياكل التشريحية الموجودة في العقد القاعدية.[20][21][22]
| التقسيم الأساسي للصفيحة القاعدية (أنبوبة عصبية) | التقسيم الثانوي | الشرائح النهائية في شخص بالغ |
|---|---|---|
| دماغ أمامي |
| |
| دماغ متوسط |
|
|
| دماغ خلفي |
| |
العقد القاعدية تشكل عنصرا أساسيا في المخ. وعلى النقيض من الطبقة القشرية التي تبطن سطح الدماغ الأمامي، فإن العقد القاعدية عبارة عن مجموعة كتل من المادة الرمادية الكامنة في أعماق المخ وهي ليست بعيدة عن المهاد. ومثل معظم أجزاء الدماغ، تتكون العقد القاعدية من الجانبين الأيمن والأيسر وهما صورتان متطابقة ظاهرتان لبعضهما البعض.[24]
من حيث التشريح، تنقسم العقد القاعدية إلى أربعة أنوية، اعتمادا على مدى قربها من قمة الرأس: النواة الذنبية (وهي أكبر نواة) والبَطامة (Putamen) والكرة الشاحبة (Globus pallidum): تتألف من جزء باطن وجزء خارجي والمادة اللامسمّاة (Substantia innominata)
في الرسم التوضيحي إلى اليمين، يُظهر الدماغ البشري ومكونات العقد القاعدية.[25][26]

تاريخ
استغرق وقتًا طويلًا حتى تم القبول أن العقد القاعدية تشكل جزءًا رئيسيًا من النظام الدماغي. نشر ثوماس ويليس في عام 1664 التشخيص التشريحي الأول لمباني تحت قشرية محددة.[27]
انظر أيضًا
مراجع
- Stocco، Andrea؛ Lebiere، Christian؛ Anderson، John R. (2010). "Conditional Routing of Information to the Cortex: A Model of the Basal Ganglia's Role in Cognitive Coordination". Psychological Review. ج. 117 ع. 2: 541–74. DOI:10.1037/a0019077. PMC:3064519. PMID:20438237.
- Weyhenmeyer، James A.؛ Gallman، Eve. A. (2007). Rapid Review of Neuroscience. Mosby Elsevier. ص. 102. ISBN:978-0-323-02261-3. مؤرشف من الأصل في 2022-05-12.
- Fix، James D. (2008). "Basal Ganglia and the Striatal Motor System". Neuroanatomy (Board Review Series) (ط. 4th). Baltimore: Wulters Kluwer & Lippincott Williams & Wilkins. ص. 274–281. ISBN:978-0-7817-7245-7. مؤرشف من الأصل في 2022-04-08.
- Cameron IG، Watanabe M، Pari G، Munoz DP (يونيو 2010). "Executive impairment in Parkinson's disease: response automaticity and task switching". Neuropsychologia. ج. 48 ع. 7: 1948–57. DOI:10.1016/j.neuropsychologia.2010.03.015. PMID:20303998. S2CID:9993548.
- Chakravarthy، V. S.؛ Joseph، Denny؛ Bapi، Raju S. (2010). "What do the basal ganglia do? A modeling perspective". Biological Cybernetics. ج. 103 ع. 3: 237–53. DOI:10.1007/s00422-010-0401-y. PMID:20644953. S2CID:853119.
- Turner, Robert S; Desmurget, Michel (1 Dec 2010). "Basal ganglia contributions to motor control: a vigorous tutor". Current Opinion in Neurobiology. Motor systems – Neurobiology of behaviourv (بالإنجليزية). 20 (6): 704–716. DOI:10.1016/j.conb.2010.08.022. ISSN:0959-4388. PMC:3025075. PMID:20850966.
- Frank, O'Reilly (2006). "A Mechanistic Account of Striatal Dopamine Function in Human Cognition: Psychopharmacological Studies With Cabergoline and Haloperidol". Behavioral Neuroscience. American Psychological Association. ج. 120 ع. 3: 497–517. DOI:10.1037/0735-7044.120.3.497. PMID:16768602.
- Inta، D.؛ Meyer-Lindenberg، A.؛ Gass، P. (2010). "Alterations in Postnatal Neurogenesis and Dopamine Dysregulation in Schizophrenia: A Hypothesis". Schizophrenia Bulletin. ج. 37 ع. 4: 674–80. DOI:10.1093/schbul/sbq134. PMC:3122276. PMID:21097511.
- Hikosaka، O؛ Takikawa، Y؛ Kawagoe، R (2000). "Role of the basal ganglia in the control of purposive saccadic eye movements". Physiological Reviews. ج. 80 ع. 3: 953–78. DOI:10.1152/physrev.2000.80.3.953. PMID:10893428. S2CID:7502211.
- Eisinger, Robert S.; Urdaneta, Morgan E.; Foote, Kelly D.; Okun, Michael S.; Gunduz, Aysegul (2018). "Non-motor Characterization of the Basal Ganglia: Evidence From Human and Non-human Primate Electrophysiology". Frontiers in Neuroscience (بالإنجليزية). 12: 385. DOI:10.3389/fnins.2018.00385. ISSN:1662-453X. PMC:6041403. PMID:30026679.
- Eisinger، RS؛ Urdaneta، ME؛ Foote، KD؛ Okun، MS؛ Gunduz، A (2018). "Non-motor Characterization of the Basal Ganglia: Evidence From Human and Non-human Primate Electrophysiology". Frontiers in Neuroscience. ج. 12: 385. DOI:10.3389/fnins.2018.00385. PMC:6041403. PMID:30026679.
- Hélie، Sébastien؛ Ell، Shawn W.؛ Ashby، F. Gregory (1 مارس 2015). "Learning robust cortico-cortical associations with the basal ganglia: an integrative review". Cortex. ج. 64: 123–135. DOI:10.1016/j.cortex.2014.10.011. ISSN:1973-8102. PMID:25461713. S2CID:17994331.
- Schroll، Henning؛ Hamker، Fred H. (30 ديسمبر 2013). "Computational models of basal-ganglia pathway functions: focus on functional neuroanatomy". Frontiers in Systems Neuroscience. ج. 7: 122. DOI:10.3389/fnsys.2013.00122. ISSN:1662-5137. PMC:3874581. PMID:24416002.
- DeLong MR، Wichmann T (يناير 2007). "Circuits and circuit disorders of the basal ganglia". Arch. Neurol. ج. 64 ع. 1: 20–4. DOI:10.1001/archneur.64.1.20. PMID:17210805. مؤرشف من الأصل في 2010-01-13.
- Kempton MJ، Salvador Z، Munafò MR، Geddes JR، Simmons A، Frangou S، Williams SC (2011). "Structural Neuroimaging Studies in Major Depressive Disorder: Meta-analysis and Comparison With Bipolar Disorder". Arch Gen Psychiatry. ج. 68 ع. 7: 675–90. DOI:10.1001/archgenpsychiatry.2011.60. PMID:21727252. مؤرشف من الأصل في 2023-01-29. see also MRI database at www.depressiondatabase.org نسخة محفوظة 13 مايو 2012 على موقع واي باك مشين.
- Radua، Joaquim؛ Mataix-Cols، David (نوفمبر 2009). "Voxel-wise meta-analysis of grey matter changes in obsessive–compulsive disorder". British Journal of Psychiatry. ج. 195 ع. 5: 393–402. DOI:10.1192/bjp.bp.108.055046. PMID:19880927.
- Radua، Joaquim؛ van den Heuvel، Odile A.؛ Surguladze، Simon؛ Mataix-Cols، David (5 يوليو 2010). "Meta-analytical comparison of voxel-based morphometry studies in obsessive-compulsive disorder vs other anxiety disorders". Archives of General Psychiatry. ج. 67 ع. 7: 701–711. DOI:10.1001/archgenpsychiatry.2010.70. PMID:20603451.
- Alm، Per A. (2004). "Stuttering and the basal ganglia circuits: a critical review of possible relations". Journal of communication disorders. ج. 37 ع. 4: 325–69. DOI:10.1016/j.jcomdis.2004.03.001. PMID:15159193.
- Marín & Rubenstein. (2001). A Long, Remarkable Journey: Tangential Migration in the Telencephalon. Nature Reviews Neuroscience, 2.
- Stocco، Andrea؛ Lebiere، Christian؛ Anderson، John R. (2010). "Conditional Routing of Information to the Cortex: A Model of the Basal Ganglia's Role in Cognitive Coordination". Psychological Review. ج. 117 ع. 2: 541–74. DOI:10.1037/a0019077. PMC:3064519. PMID:20438237.
- Fix، James D. (2008). "Basal Ganglia and the Striatal Motor System". Neuroanatomy (Board Review Series) (ط. 4th). Baltimore: Wulters Kluwer & Lippincott Wiliams & Wilkins. ص. 274–281. ISBN:0-7817-7245-1. مؤرشف من الأصل في 2022-04-08.
- Regina Bailey. "Divisions of the Brain". about.com. مؤرشف من الأصل في 2010-12-02. اطلع عليه بتاريخ 2010-11-30.
- Sano، T (1910). "Beitrag zur vergleichenden Anatomie der Substantia nigra, des Corpus Luysii und der Zona incerta". Mschr Psychiat Neurol. ج. 28 ع. 1: 26–34. DOI:10.1159/000209678.
- Hall، John (2011). Guyton and Hall textbook of medical physiology (ط. 12th). Philadelphia, Pa.: Saunders/Elsevier. ص. 690. ISBN:978-1-4160-4574-8.
- Voorn، Pieter؛ Vanderschuren، Louk J. M. J.؛ Groenewegen، Henk J.؛ Robbins، Trevor W.؛ Pennartz، Cyriel M. A. (1 أغسطس 2004). "Putting a spin on the dorsal-ventral divide of the striatum". Trends in Neurosciences. ج. 27 ع. 8: 468–474. DOI:10.1016/j.tins.2004.06.006. ISSN:0166-2236. PMID:15271494.
- Burton، AC؛ Nakamura، K؛ Roesch، MR (يناير 2015). "From ventral-medial to dorsal-lateral striatum: neural correlates of reward-guided decision-making". Neurobiology of learning and memory. ج. 117: 51–9. DOI:10.1016/j.nlm.2014.05.003. PMC:4240773. PMID:24858182.
- Andrew Gilies, A brief history of the basal ganglia, retrieved on 27 June 2005 نسخة محفوظة 23 نوفمبر 2013 على موقع واي باك مشين. "نسخة مؤرشفة". مؤرشف من الأصل في 2006-10-08. اطلع عليه بتاريخ 2018-04-30.
{{استشهاد ويب}}: صيانة الاستشهاد: BOT: original URL status unknown (link)
 بوابة تشريح
بوابة تشريح بوابة طب
بوابة طب بوابة علوم عصبية
بوابة علوم عصبية