عديد الريبوسومات
عديد الريبوسومات (بالإنجليزية: polyribosome) أو عديد الجسيمات (بالإنجليزية: polysome) هو مجموعة من الريبوسومات متصلة بجزيء واحد من الرنا المرسال كالخرز في مسبحة.[1] وهو عبارة عن مركب معقد من رنا مرسال واثنين أو أكثر من الريبوسومات التي تعمل على ترجمة تعليمات الرنا المرسال إلى عديدات الببتيد. سُميت في الأصل "إرغوسومات" في عام 1963، ولاحقاً تم وصفها أكثر من قبل جوناثان وارنر، بول م. كنوبف،[2] وأليكس ريتش.
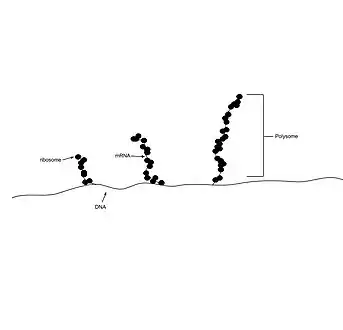
تتشكل عديدات الريبوسومات أثناء مرحلة الإطالة عندما تقوم الريبوسومات وعوامل الإطالة بتفسير عديد الببتيد المشفر. تتحرك عدة ريبوسومات على طول منطقة الترميز على الرنا المرسال، مشكلةً عديد الريبوسومات. إن قدرة عدة ريبوسومات على العمل على جزيء رنا مرسال واحد يفسر الكمية القليلة من جزيئات الرنا المرسال في الخلية.[3] تختلف بنية عديد الريبوسومات بين بدائيات النوى، حقيقيات النوى، وعديدات الجسيمات المرتبطة بأغشية الخلايا. [1] يمكن استخدام نشاط عديدات الجسيمات لقياس مستوى التعبير الجيني في الخلية من خلال تقنية تسمى تنميط عديدات الريبوسومات.[4]
البنية
كانت تقنيات المجهر الإلكتروني كالتلوين، تظليل المعادن، وقطع الخلايا الرقيقة للغاية هي الطرق الأصلية لتحديد بنية عديد الجسيمات. سمح تطور تقنيات المجهر الإلكتروني فائق البرودة بزيادة دقة الصورة، مما أدى إلى طريقة أكثر دقة لتحديد بنية عديد الجسيمات. يمكن أن تعكس البنى المختلفة من عديدات الريبوسومات مرحلة عملية ترجمة الرنا المرسال. بملاحظة تغير أشكال عديدات الريبوسومات تم العثور على عدد كبير من عديدات الريبوسومات الدائرية والمتعرجة بعد جولات قليلة من الترجمة. بعد فترة أطول من الترجمة تكونت أشكال أخرى حلزونية كثيفة ثلاثية الأبعاد.[1] تنتج الخلايا المختلفة بنى مختلفة من عديدات الجسيمات.
بدائيات النوى
لُوحظ أن عديدات الجسيمات البكتيرية تشكل بنى ثنائية الصفوف. في هذه البنية، ترتبط الريبوسومات ببعضها البعض من خلال وحيدات أصغر. عادةً ما يكون لهذه البنى المزدوجة مساراً جيبياً (متعرجاً) أو مساراً حلزونياً ثلاثيّ الأبعاد. في المسار الجيبي، هناك نوعان من الروابط بين الوحيدات الصغيرة: "من الأعلى إلى الأعلى" أو "من الأعلى إلى الأسفل". أما في المسار الحلزوني ثلاثي الأبعاد، لوحظ نوع واحد من الارتباطات "من الأعلى إلى الأعلى".[1]
توجد عديدات الجسيمات في العتائق، ولكن بنيتها غير معروفة تماماً.[5]
داخل الخلايا
أظهرت الدراسات في داخل الخلايا أن عديدات الجسيمات في حقيقيات النوى تُظهر بنى خطية. تم العثور على عديدات الجسيمات الحلزونية الكثيفة ثلاثية الأبعاد وثنائية الصفوف المستوية مع ارتباطات متغيرة ومنها "من الأعلى إلى الأعلى" المشابهة لعديدات الجسيمات في بدائيات النوى. تشبه عديدات الجسيمات ذات البنية ثلاثية الأبعاد في حقيقيات النوى مثيلاتها في بدائيات النوى في أنها عبارة عن "حلزون كثيف مع أربعة ريبوسومات في كل دور". يمكن أن تدل هذه البنية الكثيفة على وظيفتها كمنظمات للترجمة. توجد عديدات الجسيمات ثلاثية الأبعاد في خلايا الساركوما.[1]
خارج الخلايا
أظهرت الفحصوات بمجهر القوة الذرية في الدراسات المختبرية أنه يمكن تشكيل عديدات جسيمات دائرية الشكل بواسطة الرنا المرسال الحر عديد الأنين بوجود عامل البدء (eIF4E) مرتبطاً بالقلنسوة على النهاية 5' والعامل (PABP) مرتبطاً بالذيل عديد الأدنين على النهاية 3'. ومع ذلك، فإن هذا التفاعل بين القلنسوة والذيل بوساطة بروتين معقد ليس طريقة فريدة لتدوير الرنا المرسال متعدد الريبوسومات. وقد وجد أنه يمكن تشكيل عديدات الريبوسومات بنجاح أثناء عملية الترجمة بوجود رنا مرسال بلا قلنسوة أو ذيل عديد الأدنين وكذلك الأمر مع الرنا المرسال المغطى بقلنسوة ولكن بلا ذيل. [1]
أغشية الخلايا
عديدات الجسيمات المرتبطة بأغشية الخلايا لها بنية ثنائية الأبعاد نظراً لوجودها على سطح الغشاء. تؤدي هذه البنية المقيدة إلى تدويرعديدات الريبوسومات بحيث تكون الريبوسومات مرتبة مكونة مناطق دخول وخروج سلسة. يتم تدوير كل ريبوسوم بزاوية مطابقة لسابقتها، مشكلةً دوامة ثنائية الأبعاد.[1]
التنميط
تنميط عديد الريبوسومات هو تقنية تستخدم الهكساميد الحلقي لإيقاف الترجمة ومحلول السكروز لفصل المواد الناتجة بعملية التثفل.[3] تتسدم جزيئات الرنا المرسال المرتبطة بريبوسوم واحد بشكل أسرع من جزيئات الرنا المرسال الحرة وتتسدم جزيئات الرنا المرسال المرتبطة بعديد الريبوسومات بشكل أسرع من الريبوسومات المرتبطة بريبوسوم واحد. تم الكشف عن العديد من القمم المتقاللرنا المرسال من خلال قياس إجمالي البروتين في المحلول. تزداد القمم المتقابلة للرنا المرسال بازدياد نسبة الريبوسومات التي تكوّن عديدات ريبوسومات. يدل وجود الرنا المرسال في المحلول عن استمرارية عملية الترجمة. يتم تطبيق التنميط على النحو الأمثل على الخلايا والأنسجة المستنبتة لتتبع حالة عملية الترجمة للرنا المرسال وكذلك قياس كثافة الريبوسوم.[4] تم استخدام هذه التقنية لمقارنة حالات عمليات ترجمة الرنا المرسال في أنواع الخلايا المختلفة.
على سبيل المثال، تم استخدام التنميط في دراسة للتحقق من تأثير فيروس التهاب الفم الحويصلي (VSV) في خلايا الثدييات.[6] أظهرت البيانات أن عديدات الجسيمات في الرنا المرسال الخاص بالفيروسات أكثر من عديدات الجسيمات في الرنا المرسال الخاص بالخلايا المضيفة، وبالتالي تراجع ترجمة جزيئات الرنا المرسال للمضيف وزيادة ترجمة جزيئات الرنا المرسال الفيروسية.
المراجع
- "Three-Dimensional Organization of Polyribosomes- A Modern Approach". Biochemistry. Biokhimiia. ج. 83 ع. Suppl 1: S48–S55. يناير 2018. DOI:10.1134/S0006297918140055. PMID:29544430.
- Cambra، Kris (Spring 2017). "Paul M. Knopf, PhD". Brown Medicine. Brown University. مؤرشف من الأصل في 2017-09-04. اطلع عليه بتاريخ 2017-07-24.
- "Translatome profiling: methods for genome-scale analysis of mRNA translation". Briefings in Functional Genomics. ج. 15 ع. 1: 22–31. يناير 2016. DOI:10.1093/bfgp/elu045. PMID:25380596.
- "Biogenesis of phased siRNAs on membrane-bound polysomes in Arabidopsis". eLife. ج. 5: e22750. ديسمبر 2016. DOI:10.7554/eLife.22750. PMC:5207768. PMID:27938667.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة)صيانة الاستشهاد: دوي مجاني غير معلم (link) - "Transcription and translation are coupled in Archaea". Molecular Biology and Evolution. ج. 24 ع. 4: 893–5. أبريل 2007. DOI:10.1093/molbev/msm007. PMID:17237472.
- "Global analysis of polysome-associated mRNA in vesicular stomatitis virus infected cells". PLOS Pathogens. ج. 15 ع. 6: e1007875. يونيو 2019. DOI:10.1371/journal.ppat.1007875. PMC:6608984. PMID:31226162.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link)
وصلات خارجية
| مقدمة لعلم الوراثة |  | ||||
|---|---|---|---|---|---|
| النسـخ |
| ||||
| الترجمـة |
| ||||
| مراقبة علم التخلق |
| ||||
 بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة تقانة حيوية
بوابة تقانة حيوية