تطوي البروتين
تطوي البروتين هو عملية فيزيائية يتخذ فيها البروتين بنية ثلاثية الأبعاد طبيعية تمكنه من القيام بوظيفة بيولوجية محددة. في هذه العملية يتطوى عديد ببتيد من التفاف عشوائي [الإنجليزية] إلى بنية ثلاثية الأبعاد مميزة لها وظيفة خاصة،[1] يتواجد كل بروتين على شكل عديد ببتيد غير متطوي أو ملتف بشكل عشوائي حين يُترجم الرنا الرسول إلى سلسلة خطية من الأحماض الأمينية، ولا يملك عديد الببتيد هذا أي بنية مستقرة (ثابتة لمدة طويلة) ثلاثية الأبعاد. بينما يتم تخليق وتركيب سلسلة عديد الببتيد بواسطة الريبوسوم، تبدأ السلسلة بالتطوي إلى بنية ثلاثية الأبعاد.
يمكن أن يبدأ تطوي العديد من البروتينات قبل انتهاء ترجمة سلسلة عديد الببتيد، حيث تتآثر الأحماض الأمينية فيما بينها لإنتاج بنية ثلاثية الأبعاد محددة بدقة تُعرف باسم الحالة الواطنة. تُحدد هذه البنية ثلاثية الأبعاد بواسطة تسلسل وترتيب الأحماض الأمينية في سلسلة عديد الببتيد (مبدأ أنفينسن [الإنجليزية]).[2]
البنية الصحيحة ثلاثية الأبعاد ضرورية لأداء الوظيفة، رغم أن بعض أجزاء بعض البروتينات الوظيفية قد تبقى غير متطوية،[3] وضرورية لفهم حركيات البروتين [الإنجليزية]. الفشل في التطوي إلى الهيئة الطبيعية يُنتج في العادة بروتينات غير وظيفية، لكن يمكن أن يكون لدى البروتينات خاطئة التطوي في بعض الأحيان وظيفة معدلة أو سامة. يُعتقد أن العديد من أمراض التنكس العصبي وغيرها ناتجة من تراكم لييفات نشواني مكونة من بروتينات خاطئة التطوي.[4] سبب العديد من الحساسيات هو التطوي غير الصحيح لبعض البروتينات، لأن جهاز المناعة لا يُنتج أجساما مضادة لبعض بُنى البروتين.[5]
تمسخ البروتينات هو العملية العكسية، أي الانتقال من هيئة التطوي إلى هيئة عدم التطوي، وتحدث أثناء الطهي والاحتراق والاعتلالات البروتينية والسياقات الأخرى.
تختلف مدة التطوي بشكل كبير جدا حسب البروتين، حين يُدرس البروتين خارج الخلية، فإن أبطأ البروتينات في التطوي تحتاج عدة دقائق أو ساعات لتتطوى وذلك بسبب مصاوغة البرولين ويجب أن تمر عبر حالات وسيطة -مثل نقاط التحقق- قبل أن تتم العملية.[6] من ناحية أخرى، عادة ما تتطوى البروتينات أحادية النطاق الصغيرة جدا التي لا يتجاوز طولها مئة حمض أميني في خطوة واحدة.[7] يكون مقياس الزمن بالميليثانية وأسرع تفاعلات تطوي البروتين المعروفة تتم خلال عدة ميكرو ثواني.[8] يعتمد المقياس الزمني لتطوي بروتين ما على حجمه، رتبة الاتصال [الإنجليزية] وطوبولوجيا الدارة [الإنجليزية].
فهم آليات عملية تطوي البروتين ومحاكاتها هي تحديات مهمة لعلم الأحياء الحاسوبي مند عقد 1960.
عملية تطوي البروتين
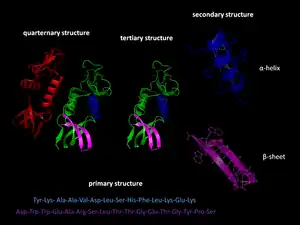
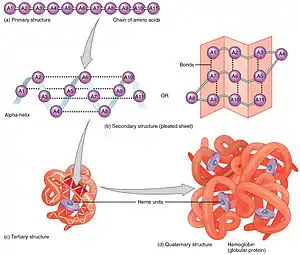
البنية الأولية
بنية البروتين الأولية هي تسلسل خطي من الأحماض الأمينية يحدد هيئة التطوي الطبيعية.[9] نوع الأحماض الأمينية ومواضعها في عديد الببتيد هي العوامل التي تحدد ماهي أجزاء البروتين التي ستتطوى قرب بعضها لتشكيل البنية ثلاثية الأبعاد. تركيب الأحماض الأمينية ليس مهما بقدر تسلسلها.[10] الحقيقة الأساسية حول التطوي هي أن تسلسل الأحماض الأمينية لكل بروتين يحتوي المعلومة التي تحدد البنية الطبيعية والمسار الذي يؤدي إلى تلك الحالة. لكن هذا لا يعني أن تسلسلات الأحماض الأمينية شبه المتماثلة تتطوى دائما بنفس الطريقة.[11] تختلف هيئات التطوي بناء على عوامل بيئية كذلك، حيث تتطوى بروتينات متماثلة بشكل مختلف حسب المكان الذي تتواجد فيه.
البنية الثانوية
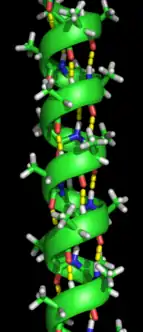
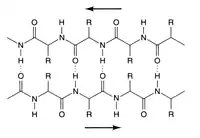
تشكيل البنية الثانوية هي الخطوة الأولى في عملية التطوي التي يقوم بها البروتين ليتخذ هيئته الطبيعية. من خصائص تكون البنية الثانوية وجود البُنى الثانوية: لوالب ألفا وصحائف بيتا التي تتطوى بسرعة لأنها تستقر بواسطة روابط هيدروجينية داخل جزيئية، وهذا ما وضحه لينوس باولنغ أولا. يوفر تكوُّن الروابط الهيدروجينية داخل الجزيئية عاملا آخرا مهما في استقرار بنية البروتين.[12] تتشكل لوالب ألفا بواسطة روابط هيدروجينية بين العمود الفقري لتكوين شكل لولبي (الصورة على اليسار).[10] صحيفة بيتا المطوية هي بنية أقل تواترا من لوالب ألفا وتتكون عبر انطواء العمود الفقري على نفسه لتشكيل روابط هيدروجينية (كما هو موضح في الشكل على اليمين).
تتكون الروابط الهيدروجينية بين هيدروجين الأميد وأكسجين الكربونيل الخاصين بالرابطة الببتيدية. توجد صحائف بيتا متضادة التوازي وكذلك صحائف بيتا المتوازية ويكون استقرار روابط الهيدروجين أقوى في صحائف بيتا متضادة التوازي لأن روابط الهيدروجين فيها بزاوية مثالية قدرها 180 درجة مقارنة بروابط الهيدروجين المائلة الخاصة بصحائف بيتا المتوازية.[10]
البنية الثالثية
يمكن أن تكون لوالب ألفا وصحائف بيتا مزدوج الألفة في طبيعتها، أو تحتوي على جزء محب للماء وآخر كاره له. تساعد هذه الخصائص الخاصة بالبنى الثانوية على تشكل البنية الثالثية للبروتين، بحيث يحدث التطوي بطريقة تقابل فيها الأجزاء المحبة للماء البيئة المائية المحيطة بالبروتين أما الأجزاء الكارهة للماء فتتجه صوب لب البروتين الكاره للماء.[13] يسمح التنظيم الهرمي للبنى الثانوية بتشكل البنية الثالثية النهائية، وعند تشكل هذه الأخيرة واستقرارها بواسطة التآثرات الكارهة للماء، يمكن أن تكون هنالك روابط تساهمية على شكل جسور ثنائي كبريتيد بين وحدتي سيستئين. البنية الثالثية لبروتين هي تطوي سلسلة عديد ببتيد واحدة فقط، لكن يمكن لتآثرات إضافية بين عدة سلاسل عديد ببتيد أن تُشكل بنية رابعية.[14]
البنية الرابعية
يمكن أن تُشكل العديد من البنى الثالثية بنية بروتين رابعية، والتي عادة ما يتم فيها «تجميع» أو «التجميع المشترك» لعدة وحدات فرعية متطوية مسبقا، بعبارة أخرى يمكن أن تتآثر عدة سلاسل عديد ببتيد لتشكل بروتينا رابعيا وظيفيا بالكامل[10] يسمى في العادة معقد بروتيني.
القوى المسببة لتطوي البروتين
التطوي هو عملية تلقائية [الإنجليزية] تقودها بشكل أساسي: التآثرات الكارهة للماء، تكون الروابط الهيدروجينية داخل الجزيئية، قوى فان دير فالس، وتعاكسها الإنتروبيا الهيئية [الإنجليزية].[15] عادة ما تبدأ عملية التطوي أثناء الترجمة، حيث تبدأ النهاية الأمينية بالتطوي بينما لا يزال جزء النهاية الكربوكسيلية في طور التخليق بواسطة الريبوسوم، ومع ذلك يمكن أن يتطوى جزيء البروتين تلقائيا أثناء أو بعد التخليق الحيوي.[16] في حين يُعتبر أن هذه الجزيئات الضخمة تقوم "بطي نفسها"، تعتمد العملية كذلك على: المذيب (ماء أو ليبيد ثنائي الطبقة)،[17] تركيز الأملاح، الأس الهيدروجيني، درجة الحرارة، واحتمال تواجد العوامل المرافقة والشابرونات،
للبروتينات حدود في إمكانية تطويها وذلك حسب زوايا الانحناء المقيدة أو الهيئات الممكنة. توصف وتُرسم هذه الزوايا السامحة لتطوي البروتين التي يمكنه تحقيقها أثناء التطوي بمخطط ثنائي الأبعاد يعرف بمخطط راماشاندرن [الإنجليزية] حيث توضح بزاويتي بسي (ψ) وفي (Φ) اللتان تحددان مدى التطوي المسموح به طاقويا الذي لا تمنعه تآثرات الذرات فيما بينها.[18]
تأثير كره الماء
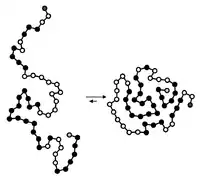
يجب أن يكون تطوي البروتين مفضلا ثرموديناميكيا داخل الخلايا في سبيل أن يكون تلقائيا. ولأنه من المعروف أن تطوي البروتين تفاعل تلقائيٌ، يجب أن تكون له قيمة سالبة لطاقة غيبس الحرة، وطاقة غيبس الحرة مرتبطة بشكل وثيق مع الإنثالبيا والإنتروبيا [10] (تُعطى بالمعادلة ). لكي يكون لتطوي البروتين قيمة دلتا G سالبة ويكون مفضلا ثرموديناميكيا يجب أن تكون الإنثالبيا أو الإنتروبيا مفضلة أو كلاهما.

تخفيض عدد السلاسل الجانبية المتعرضة للماء قوة مهمة مسببة لعملية التطوي.[19] تأثير كره الماء هي ظاهرة تتطوى فيها سلاسل البروتين الكارهة للماء نحو لب البروتين (بعيدا عن المحيط المحب للماء).[10] في البيئة المائية، تميل جزيئات الماء للتجمع حول المناطق أو السلاسل الجانبية الكارهة للماء، مشكلة صدفات أو أغلفة مكونة من جزيئات مائية منتظمة.[20] يزيد انتظام جزيئات الماء حول منطقةٍ كارهةٍ للماء من انتظام النظام وبذلك يساهم في تغير سلبي في الإنتروبيا (إنتروبيا أقل في النظام). يؤدي انتظام جزيئات الماء في هذه الأغلفة المائية إلى انهيار كره الماء [الإنجليزية] أو تطوي المجموعات الكارهة للماء نحو الداخل. يُعيد انهيار كره الماء الإنروبيا إلى النظام عبر كسر الأغلفة المائية الذي يحرر ويزيل انتظام جزيئات الماء.[10] يساهم تعدد المجموعات الكارهة للماء المتآثرة داخل لب البروتين المتطوي كرويا بنسبة معتبرة من استقرار بنية البروتين بعد التطوي، بسبب التراكم الكبير لقوى فان دير فالس (بالتحديد قوى تشتت لندن).[10] يكون تأثير كره الماء قوة مسببة للتطوي ثرموديناميكيا فقط عند تواجد وسط مائي فيه جزيء مزدوج الألفة يحتوي منطقة كبيرة كارهة للماء.[21] تعتمد قوة الروابط الهيدروجينية على بيئتها، فالروابط الموجودة داخل لب كاره للماء تساهم بقوة أكثر في استقرار هيئة التطوي الطبيعية من الروابط المتعرضة لبيئة مائية.[22]
في البروتينات ذات التطويات الكروية، تميل الأحماض الأمينية الكارهة للماء لأن تتخلل التسلسل الأولي بانتظام على طوله، بدل أن تتوزع عشوائيا أو تتكدس معا في منطقة ما.[23][24] مع ذلك، تُظهر البروتينات الجديدة [الإنجليزية] التي نشأت حديثا والتي تميل لأن تكون مضطربة ذاتيا[25][26] نمطا معاكسا يحدث فيه تكدس للأحماض الأمينية الكارهة للماء على طول التسلسل الأولي.[27]
الشابرونات
الشابرونات هي قسم من البروتينات التي تساعد في التطوي الصحيح للبروتينات حيويا. تتواجد الشابرونات في جميع الأحياز الخلوية وتتآثر مع سلسلة عديد الببتيد من أجل السماح للهيئة البروتين الطبيعية ثلاثية الأبعاد بالتشكل، لكن الشابرونات في حد ذاتها ليست جزءا من بنية البروتين النهائية التي تساعد في تطويها.[28] يمكن أن تساعد الشابرونات حتى في تطوي عديد الببتيد الناشئ الذي يتم تخليقه بواسطة الريبوسوم.[29] تعمل الشابرونات عبر الارتباط ببنية بروتين غير مستقرة وتحقيق استقرارها في مسار تطويها. لا تملك الشابرونات المعلومات الضرورية لمعرفة هيئة التطوي الطبيعية الخاصة بالبروتين الذي تساعده وإنما تعمل على منع هيئات التطوي غير الصحيحة.[29] وهذا يعني أن الشابرونات لا تزيد سرعة الخطوات الفردية في مسار التطوي إلى الهيئة الطبيعية بل تعمل على تخفيض التطويات الممكنة غير المرغوب فيها والتي -من دون الشابرونات- كانت ستُبطئ البحث عن هيئة وسطية مناسبة، وتوفر مسارا أكثر كفاءة لسلاسل عديد الببتيد كي تتخذ الهيئات الصحيحة.[28] لا يجب الخلط بين الشابرونات ومحفزات التطوي، تقوم هذه الأخيرة فعلا بتحفيز مراحل -تعتبر بطيئة من دون تدخلها- في مسار التطوي. من الأمثلة على محفزات التطوي مصاوغ ثنائي كبريتيد البروتين [الإنجليزية] ومصاوغ البروليل اللذان يمكن أن يكون لهما دور في تكوين روابط ثنائي كبريتيد أو التحويل بين المصاوغات الفراغية سيس وترانس، على التوالي.[29]
الشابرونات مهمة لتطوي البروتين حيويا لأنها توفر الدعم اللازم للبروتين ليتخذ ترتيباته وهيئاته الصحيحة بفعالية كافية لدرجة يُعتبر أن دورها «ذو صلة بيولوجيا».[30] وهذا يعني أن بإمكان سلسلة عديد الببتيد نظريا التطوي إلى بنيتها الطبيعية من دون مساعدة الشابرونات، كما تم إثباته بواسطة تجارب تطوي البروتين في المختبر.[30] لكن ثبت أن تلك العملية غير فعالة للغاية أو بطيئة للغاية لكي تتواجد في الأنظمة البيولوجية، لذلك الشابرونات ضرورية لتطوي البروتين حيويا. بالإضافة إلى دورها في المساعدة على تشكل البنية الطبيعية، تقوم الشابرونات بعدة وظائف أخرى مثل: نقل البروتين، تفكيك البروتين، وحتى السماح للبروتينات المتمسخة المتعرضة لبعض عوامل التمسيخ الخارجية بإعادة التطوي إلى بٌناها الطبيعية الصحيحة.[31]
لا يملك البروتين المتمسخ كليا كلا البنيتين الثالثية والثانوية، ويتواجد على شكل التفاف عشوائي [الإنجليزية]. يمكن لبعض البروتينات تحت شروط معينة أن تعاود التطوي، لكن في أغلب الحالات يكون التسمخ غير قابل للعكس.[32] في بعض الأحيان تحمي الخلايا بروتيناتها من التمسخ بسبب تأثير الحرارة المرتفعة عبر إنزيمات تُعرف ببروتينات الصدمة الحرارية (نوع من الشابرونات) والتي تساعد بروتينات أخرى في التطوي والبقاء متطوية. توجد بروتينات الصدمة الحرارية في جميع الأنواع التي تم اختبارها من البكتيريا إلى البشر، وهذا يوحي بأنها تطورت في البدايات الأولى وأن لديها دورا مهما. بعض البروتينات لا تتطوى في الخلية نهائيا إلا بمساعدة الشابرونات التي إما تعزل البروتينات بشكل منفرد لكي لا يُقاطَع تطويها بتآثراتٍ مع بروتينات أخرى وإما تساعد في فك تطوي البروتينات خاطئة التطوي والسماح لها بالتطوي مجددا إلى البنية الطبيعية الصحيحة.[33] هذه الوظيفة حاسمة لمنع خطر الترسب إلى تكدسات لابلورية غير قابلة للذوبان. من العوامل الخارجية التي لها دور في تمسخ البروتين أو الإخلال ببنيته الطبيعية: الحرارة، المجالات الخارجية (كهربائية، مغناطيسية)،[34] ازدحام الجزيئات،[35] وحتى محدودية الحيز الخلوي الذي يمكن أن يكون له تأثير كبير على التطوي،[36] التراكيز العالية للمذاب، الأس الهيدروجيني العالي، القوى الميكانيكية، تواجد عوامل كيميائية ممسِّخة يمكن أن يساهم في تمسخ البروتين كذلك. وتُصنف هذه العوامل فرديا على أنها عوامل إجهاد أو ضغط. تتواجد الشابرونات بتراكيز متزايدة أثناء أوقات الإجهاد الخلوي للمساعدة في التطوي الصحيح للبروتينات الناشئة وكذلك إعادة تطوي البروتينات خاطئة التطوي.[28]
تحت بعض الشروط لن تتطوى البروتينات إلى هيئاتها البيوكيميائية الوظيفية. تسبب درجات الحرارة فوق أو أسفل المجال الذي تميل الخلايا للعيش فيه فك تطوي أو تمسخ البروتينات غير المستقرة حراريا [الإنجليزية] (لهذا يجعل الغليان بياض البيض غير شفاف). الاستقرار الحراري غير ثابت، فعلى سبيل المثال: البكتيريا فائقة الألفة للحرارة [الإنجليزية] يمكن أن تنمو في درجات حرارة تصل حتى 122 مئوية،[37] وهو ما يتطلب بالتأكيد أن تكون كامل بروتيناتها الحيوية وتجميعات البروتين مستقرة في تلك الدرجة أو أعلى.
بكتيريا الإشريكية القولونية هي مضيف للعاثية T4، وتشفر العاثية البروتين gp31 (P17313) الذي يظهر بأنه نظير بنيوي ووظيفي للشابرون GroES [الإنجليزية] الخاص بالإشريكية ويمكن للأخير أن يحل مكانه أثناء تجميع الفيروس عند التكاثر.[38] مثل GroES، يشكل البروتين السكري 31 معقدا مستقرا مع الشابرون GroEL [الإنجليزية] الضروري للغاية من أجل التطوي والتجميع الحيوي لبروتين قفيصة الفيروس T4 الكبير gp23.[38]
تبديل التطوي
لدى بعض البروتينات عدة هيئات طبيعية وتغير تطويها حسب عوامل خارجية، على سبيل المثال: يُغير بروتين KaiB [الإنجليزية] تطويه خلال اليوم، ويعمل كساعة للبكتيريا الزرقاء. يقدر أن حوالي 0.5-4% من بروتينات بنك بيانات البروتينات تغير تطويها أو تطوي أجزاء منها أثناء الارتباط.[39]
التطوي الخاطئ للبروتين وأمراض التحلل العصبي
يُعتبر البروتين خاطئ التطوي (سيء التطوي) إذا لم يستطع اتخاذ بنيته الطبيعية، ويمكن أن يحدث ذلك بسبب طفراتٍ في تسلسل الأحماض الأمينية أو إخلالٍ لعملية التطوي الطبيعية بواسطة عوامل خارجية.[40] تحتوي البروتينات خاطئة التطوي عادة على صحائف بيتا مرتبة على نسقٍ لتجميع الجزيئات الفائقة يعرف ببنية تصالب بيتا (cross-β structure) وهو نسق تكون فيه السلاسل الفردية لكل صحيفة بيتا متجهة عموديا بالنسبة لمحور لييف النشواني مع 4.7 أنغستروم فراغ بينها في حين تكون صحائف بيتا موازية للمحور مع 10 أنغستروم فراغ بينها.[41] التجميعات الغنية بصحائف بيتا مستقرة جدا، وغير قابلة للذوبان وهي في العادة مقاومة للتحلل البروتيني.[42] يحدث الاستقرار البنيوي لهذه التجميعات اللييفية بسبب تآثرات كثيفة بين موحودات البروتين، والتي تحدث بسبب روابط هيدروجينية بين العمود الفقري لسلاسل بيتا.[42] يمكن أن تثير البروتينات خاطئة التطوي المزيد من سوء التطوي والتراكم لبروتينات أخرى لتشكل قليلات قسيمات وتكدسات. تؤدي زيادة مستوى البروتينات المتكدسة إلى تشكل بنيات تشبه النشواني والتي يمكن أن تسبب أمراضا تنكسية وموت الخلية.[40] النشوانيات هي بُنى لييفية تحتوي على روابط هيدروجينية بين جزيئية وهي غير قابلة للذوبان بشدة وتتكون من تكدسات بروتينات خاطئة التطوي.[40] ولهذا ربما لا يتمكن لمسار البروتيزوم من تفكيك البروتينات خاطئة التطوي بفعالية كافية قبل تراكمها. يمكن للبروتينات خاطئة التطوي أن تتفاعل مع بعضها البعض وتكوِّن تكدسات منتظمة البنية وتصبح سامة عبر تآثرات بين جزيئية.[40]
البروتينات المتكدسة مرتبطة بالأمراض ذات الصلة بالبريون مثل: مرض كروتزفيلد جاكوب والتهاب الدماغ الإسفنجي البقري (جنون البقر)، وكذلك بالأمراض ذات الصلة بالنشواني مثل: مرض آلزهايمر، اعتلال عضلة القلب النشواني العائلي أو اعتلال الأعصاب النشواني العائلي،[43] وكذلك التكدسات داخل الخلوية مثل داء هنتنغتون ومرض باركنسون.[4][44] أمراض التحلل العصبي هذه التي يتعلق بدءها بالعمر مرتبطة بتكدس بروتينات خاطئة التطوي في تكدسات خارج خلوية غير قابلة للذوبان و/أو شوائب داخل خلوية منها لييفات نشواني تصالب بيتا. من غير الواضح كليا إن كانت هذه التكدسات هي السبب أو مجرد عَرَضٍ لخسارة استتباب البروتين، وهو التوازن بين تخليق وتطوي وتفكيك البروتين (دورة البروتين [الإنجليزية]). وافقت وكالة الأدوية الأوروبية على استخدام تفامديس [الإنجليزية] أو فينداكيل (مثبت حركي للترانسثيريتين [الإنجليزية] رباعي القسيمات) لعلاج أمراض نشواني ترانسثيريتين. هذا يوحي بأن عملية تشكل لييف النشواني (وليس اللييفات في حد ذاتها) هي التي تسبب تحلل نسيج ما بعد الانقسام المتساوي في أمراض النشواني البشرية.[45] سوء تطوي البروتينات وتحليلها بدل إعادة تطويها واكتسابها لوظيفتها يؤدي إلى أمراض بروتينية مثل النفاخ المرتبط بألفا 1-أنتيتريبسين، التليف الكيسي وأمراض اختزان اليحلولات حيث فقدان الوظيفة البروتينية هو سبب هذه الاضطرابات. في حين أن العلاج بتبديل البروتين كان يُستخدم تاريخيا لعلاج الاضطرابات السالف ذكرها، يَستخدم نهج مستجد شابرونات صيدلية [الإنجليزية] لطي البروتينات الطافرة لجعلها وظيفية.
تقنيات تجريبية لدراسة تطوي البروتين
يمكن الوصول إلى استنتاجات حول تطوي البروتين عبر دراسات التطفر، وعادة ما تعتمد التقنيات التجريبية المستخدمة في دراسة تطوي البروتين على التطوي أو فك التطوي التدريجي للبروتينات وملاحظة التغيرات الهيئية باستخدام تقنيات قياسية ليست من تقنيات علم البلورات.
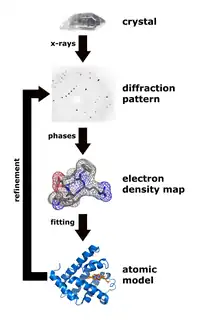
علم البلورات السيني
تقنية علم البلورات السيني هي أحد أهم الطرق الفعالة في محاولة معرفة الهيئة ثلاثية الأبعاد الخاصة ببروتين متطوي.[46] ولتنفيذ هذه التقنية، يجب وضع البروتين المراد معرفة بنيته داخل شبيكة كريستالية، ولفعل ذلك يجب أن يتوفر مذيب مناسب لعملية البلورة (طالع بالإنجليزية عملية البلورة)، والحصول على بروتين نقي مفرط التشبع في المحلول، وترسيب البلورات في المحلول.[47] بعد بلورة البروتين، يتم تسليط حزم من الأشعة السينية على الشبيكة البلورية والتي تُحيد هذه الأشعة أو تحررها خارجة منها في اتجاهات مختلفة. للأشعة الخارجة علاقة بالهيئة ثلاثية الأبعاد الخاصة بالبروتين داخل الشبيكة، وتتآثر الأشعة السينية على وجه التحديد مع سحابات الإلكترون المحيطة بالذرات المفردة داخل شبيكة البروتين وتُنتج نمط حيود قابل للتميز.[13] وفقط عبر ربط سحب كثافة الألكترون مع سعة الاشعة السينية يمكن قراءة هذه الأنماط والحصول على افتراضات حول الأطوار أو زوايا الطور التي لها دور في تحديد البنية وهو ما يجعل هذه الطريقة معقدة.[48] من دون العلاقة التي تُثبَت عبر عملية رياضية تعرف بتحويل فورييه، تجعل مشكلة الطور [الإنجليزية] التنبؤ بأنماط الحيود صعبا جدا.[13] تستخدم طرق مستجدة مثل استبدال المتشاكل المتعدد [الإنجليزية] تواجد أيون فلز ثقيل لتشتيت الأشعة السينية إلى نسق أكثر قابلية للتنبؤ، مخفضة عدد المتغيرات وبذلك تحل مشكلة الطور.[46]
مطيافية الفلورية
مطيافية الفلورية [الإنجليزية] هي طريقة حساسة للغاية لدراسة هيئة التطوي الخاصة بالبروتينات. لدى ثلاث أحماض أمينية: فينيل ألانين (Phe)، تيروسين (Tyr) وتريبتوفان (Trp) خصائص فلورية ذاتية، لكن لا يُستخدم سوى التيروسين والتريبتوفان تجريبيا وذلك بسبب مردودهما الكمي [الإنجليزية] المرتفع كفاية ليعطي إشارات فلورية جيدة. يُستثار كل من التيروسين والتريبتوفان بطول موجة قدره 280 نانومتر، في حين يستثار التريبتوفان وحده بطول الموجة 295 نانومتر. وبسبب خصائصهما العطرية غالبا ما يتواجد التريبتوفان والتيروسين مدفونان كليا أو جزئيا في الجزء الكاره للماء، في الواجهة بين نطاقي بروتين أو في الواجهة بين وحدتين فرعيتين لبروتين قليل القسيمات. في بيئة غير قطبية، يملك هذان الحمضان مردودا كيميائيا عاليا وبالتالي شِدّة فلورية عالية، وعند الإخلال ببنية البروتين الرابعية أو الثالثية، تُصبح هذه السلاسل الجانبية معرضة أكثر للبيئة المحبة للماء الخاصة بالمذيب وينخفض مردودهما الكمي وهو ما يؤدي إلى شدة فلورية منخفضة. بالنسبة لوحدات التريبتوفان يعتمد طول موجة الانبعاث الفلوري الأقصى كذلك على البيئة الموجود فيها.
يمكن أن تُستخدم مطيافية الفلورية لتحديد خصائص توازن فك التطوي [الإنجليزية] للبروتينات عبر قياس التغير في شدة انبعاث الفلورية أو طول موجة الإنبعاث الأقصى كدوال لقيمة ممسخ.[49][50] يمكن أن يكون المُمَسِّخ جزيئا كيميائيا (يوريا، كلوريد الغوانيدينيوم)، الحرارة، الأس الهيدروجيني، الضغط...إلخ. تعتمد حالة التوازن بين هيئات البروتين المختلفة والمنفصلة (هيئة طبيعية، هيئات وسطية، هيئة فك التطوي) على قيمة الممسخ، وبالتالي تعتمد إشارة الفلورية العامة الخاصة بتوازن خليط تلك الهيئات على هذه القيمة كذلك. لذلك يمكن الحصول على بروفايل يخص إشارة البروتين الفلورية العامة حسب قيمة الممسخ. يمكن أن يسمح بروفايل توازن فك التطوي بتحديد وتعريف هيئات التطوي الوسطية التي تسبق فك التطوي.[51][52] تم تطوير معادلات عامة من قبل هيوغز بيدويل للحصول على وسائط ثرموديناميكية تحدد خصائص توازن فك التطوي لبروتينات متماثلة ومتغايرة القسيمات -ثلاثية القسيمات وربما رباعية القسيمات- من هذه البروفايلات.[49] يمكن أن تُستخدم مطيافية الفلورية إلى جانب أجهزة سريعة الخلط مثل توقيف التدفق [الإنجليزية] لقياس حركيات تطوي البروتين،[53] وتوليد لطخة شيفرون [الإنجليزية] والقيام بدراسة قيمة في [الإنجليزية].
ازدواج اللون الدائري
ازدواج اللون الدائري هو أحد الوسائل العامة والأساسية في دراسة تطوي البروتين. تقيس مطيافية ازدواج اللون الدائري امتصاص الضوء المستقطب دائريا. بعض البُنى في البروتينات مثل لوالب ألفا وصحائف بيتا هي كيرالية لذلك تمتص مثل ذلك الضوء. يعمل امتصاص هذا الضوء كعلامة على درجة انطواء مجمل البروتين، واستخدمت هذه التقنية لقياس توازن فك التطوي الخاص بالبروتين عبر قياس التغير في هذا الامتصاص كدالة على تركيز المُمسخ أو الحرارة. يقيس الذوبان بالمُمسخ طاقة فك التطوي الحرة وكذلك القيمة (Cm) الخاصة بالبروتين . يقيس الذوبان بالحرارة درجة حرارة التمسخ [الإنجليزية] (Tm) بالخاصة بالبروتين.[49] كما هو الحال بالنسبة لمطيافية الفلورية، يمكن أن تُستخدم مطيافية ازدواج اللون الدائري إلى جانب أجهزة سريعة الخلط مثل توقيف التدفق [الإنجليزية] لقياس حركيات تطوي البروتين، وتوليد لطخة شيفرون [الإنجليزية].
ازدواج اللون الدائري الاهتزازي للبروتينات
توفر التقدمات الحديثة في تقنيات ازدواج اللون الدائري الاهتزازي (VCD) التي تستخدم تحويل فورييه وسائل أقوى لتحديد هيئات البروتين في المحلول، حتى بالنسبة للبروتينات الكبيرة والمعقدات. يُمكن أن تُستخدم دراسات ازدواج اللون الدائري الاهتزازي في توليفة مع بيانات حيود الأشعة السينية، وبيانات مطيافية الأشعة تحت الحمراء باستخدام تحويل فورييه (FTIR) لمحاليل البروتينات في الماء الثقيل (D2O)، أو مع الحوسبة الكمومية.
مطيافية الرنين المغناطيسي النووي للبروتين
تستطيع تقنية الرنين المغناطيسي النووي للبروتين جمع بيانات بنيوية حول البروتين عبر إحداث مجال مغناطيسي في عينات من تراكيز البروتين. في الرنين المغناطيسي النووي وحسب البيئة الكيميائية، ستمتص أنوية معينة ترددات راديوية محددة.[54][55] ولأن التغيرات البنيوية للبروتين تحدث في مقياس زمني يتراوح بين النانو ثانية والميلي ثانية، فإن تقنية الرنين المغناطيسي النووي مناسبة لدراسة تغيرات البُنى والهيئات الوسطية التي تحدث في مقاييس زمنية بين البيكو ثانية والثانية.[56] تشمل بعض طرق الرنين المغناطيسي النووي الرئيسية المستخدمة في دراسة بنية البروتينات والتغيرات البنيوية للبروتينات غير المتطوية: COSY [الإنجليزية]، وTOCSY [الإنجليزية] وHSQC [الإنجليزية] وارتخاء الزمن [الإنجليزية] (T1 وT2) وNOE [الإنجليزية].[54] NOE مفيدة بشكل خاص لأنه يمكن بواسطتها رؤية انتقالات المغنطة بين ذرات الهيدروجين المتجاورة في الفضاء.[54] تملك مختلف طرق وتجارب الرنيني المغناطيسي النووي مقاييس زمنية مختلفة تناسب كل واحدة منها تغيرات مختلفة في بنية بروتين (الصورة). يمكن لطريقة NOE التقاط (كشف) اهتزازات رابطةٍ أو استدارات سلسلة جانبية، مع ذلك هذه الطريقة حساسة أكثر من اللازم لالتقاط تطوي البروتين لأنه يحدث في مقياس زمني أكبر.[56]

لأن تطوي البروتين يحدث بين 3x 10−3 إلى 5x10−1 ثانية، أصبحت تقنيتي تشتت ارتخاء CPMG وانتقال تشبع التبادل الكيميائي [الإنجليزية] (CEST) إحدى تقنيات الرنين المغناطيسي النووي الرئيسية لدراسة التطوي.[55] فضلا عن ذلك، تُستخدم كلا التقنيتين للكشف عن الحالات (الهيئات) الوسيطة المستثارة في المسار العام لتطوي البروتين.[57] ولفعل ذلك تستغل تقنية تشتت ارتخاء CPMG ظاهرة صدى اللف، ويتم في هذه التقنية تعريض الأنوية المستهدفة إلى نبض بزاوية 90 يتبعه نبض أو عدة نبضات بزاوية 180.[58] وبينما تستعيد الأنوية ثباتها، يُشير اضطراب عريض إلى أن الأنوية المستهدفة مشاركة في الحالة الوسطية المستثارة. عبر النظر إلى مخططات تشتت الارتخاء -الناتجة- من البيانات المجمعة حول الثرموديناميكيات والحركيات بين الحالة المستثارة والحالة القاعية.[57][58] تقيس تقنية انتقال التشبع التغيرات في الإشارة من الحالة القاعية حينما تصبح الحالات المستثارة مضطربة. وتستخدم تردد تشعيع راديوي ضعيف لإشباع الحالة المستثارة لأنوية محددة والتي تنقل تشبعها إلى الحالة القاعية.[55] يتم تضخيم هذه الإشارة عبر تخفيض مغنطة (وإشارة) الحالة القاعية.[55][57]
حدود تقنية الرنين المغناطيسي النووي تكمن في أن دقتها تنخفض مع البروتينات الأكبر من 25 كيلو دالتون وأنها لا تعطي تفاصيل مثل علم البلورات السيني.[55] فضلا عن ذلك، تحليل وتفسير الرنين المغناطيسي النووي للبروتين صعب ويمكن أن يُعطي عدة حلول من نفس طيف الرنين المغناطيسي النووي.[54]
في دراسة ركزت على تطوي البروتين SOD1 [الإنجليزية] الذي له دور في التصلب الجانبي الضموري، دُرست هيئات تطوي وسطية مستثارة بواسطة تقنيتي تشتت الارتخاء وانتقال التشبع.[59] تم ربط SOD1 سابقا بعدة نسخ متطفرة مسببة للأمراض والتي افتُرض أن لها دور في تراكم البروتين، ومع ذلك ما زالت الآلية غير معروفة. باستخدام تجارب تشتت الاسترخاء وانتقال التشبع، تم اكتشاف أن عدة هيئات وسطية مستثارة تتطوى بشكل خاطئ في نسخ SOD1 الطافرة.[59]
قياس التداخل ثنائي الاستقطاب
قياس التداخل ثنائي الاستقطاب هو تقنية تعتمد على دراسة السطح وتقيس الخصائص الضوئية للطبقات الجزيئية. حين تُستخدم لتحديد خصائص تطوي البروتين، يتم بواسطتها قياس الهيئات البنيوية عبر تحديد الحجم الإجمالي لطبقة بروتين مفردة وكثافتها في الوقت الحقيقي بدقة أقل من الأنغستروم،[60] رغم أن قياسات الزمن الحقيقي لحركيات تطوي البروتين محدودة في العمليات الأبطأ من 10 هرتز. كما هو الحال في ازدواج اللون الدائري، يمكن أن يكون محفز التطوي عاملا ممسخا أو الحرارة.
دراسة التطوي بدقة زمنية عالية
تقدمت دراسة تطوي البروتين بشكل كبير في الأعوام الأخيرة بواسطة تطوير تقنيات سريعة متعلقة بالزمن [الإنجليزية]، ويتم في التجارب الخاصة بها الإثارة السريعة لتطوي عينة من بروتين غير متطوي وملاحظة الحركيات الناتجة عن ذلك. من التقنيات السريعة: تشتت النيوترون،[61] المزج فائق السرعة للمحاليل، التقنيات الضوئية الكيميائية، مطيافية زيادة الحرارة الليزرية [الإنجليزية]. من العلماء الذين ساهموا في تطوير هذه التقنيات: جيريمي كوك، هاينريش رودر، هاري بي. غراي، مارتن غروبيل، بريان داير، ويليام إيتون، شينا رادفورد، كريس دوبسون، آلان فرشت، بينغت نولتينغ ولارس كونرمان.
تحلل البروتين
يُستخدم تحلل البروتين بشكل دوري لسبر الأجزاء غير المتطوية تحت شروط محاليل مختلفة (مثل: تحلل البروتين المتوازي السريع).[62][63]
مطيافية قوة الجزيء المفرد
تُستخدم تقنيات الجزيء المفرد مثل الملاقط الضوئية وAFM لفهم آليات التطوي للبروتينات المتطوية لوحدها وكذلك للبروتينات المتطوية بمساعدة الشابرونات.[64] تُستخدم الملاقط الضوئية [الإنجليزية] لتمطيط بروتين مفرد من نهايته الآمينية أو الكربوكسيلية وإزالة تطويه للتمكن من دراسة إعادة التطوي التي تحدث بعد تمطيطه.[65] وتسمح هذه التقنية بقياس معدلات التطوي على مستوى الجزيء المفرد، على سبيل المثال: تم استخدام الملاقط الضوئية مؤخرا في دراسة تطوي وفك تطوي البروتينات التي لها دور في تخثر الدم. عامل فون ويل براند (vWF) هو بروتين له دور أساسي في علمية تكون جلطة الدم، وتم -باستخدام قياس الملاقط الضوئية مفردة الجزيء- اكتشاف أن بروتين vWF المرتبط بالكالسيوم يعمل كمستشعر لقوة القص في الدم، وتؤدي قوة القص إلى فك تطوي النطاق A2 الخاص بـvWF، والذي يتحسن معدل إعادة تطويه بشكل كبير عند تواجد الكالسيوم.[66] مؤخرا، تم توضيح أن نطاق SH3 [الإنجليزية] الخاص ببروتين src يمكن أن يدخل في عدة مسارات فك تطوي عند الخضوع لقوة.[67]
الربط بالبيوتين
يسمح الربط بالبيوتين [الإنجليزية] بأخذ لقطات تحت شروط خلوية محددة لبروتينات متطوية وغير متطوية، يُظهر الارتباط بالبيوتين تحيزا نحو البروتينات المضطربة الذاتيا المتنبئ بها.
دراسات حاسوبية لتطوي البروتين
تشمل الدراسات الحاسوبية لتطوي البروتين ثلاث نواحٍ رئيسية تتعلق بالتنبؤ باستقرارية البروتين، حركياته، وبنيته. تلخص مراجعة حديثة طرق الحوسبة المتوفرة لدراسة تطوي البروتين.[68]
مفارقة ليفينثال
أشار سايروس ليفينثال في عام 1969 إلى أنه بسب العدد الكبير جدا لدرجات الحرية في سلسلة عديد ببتيد غير متطوية، فإن الجزيء لديه عدد فلكي من الهيئات البنيوية التي يمكن أن يتخذها، وقدَّر في أحد أوراقه البحثية العدد بـ3300 أو 10143.[69] مفارقة ليفينثال [الإنجليزية] هي تجربة فكرية مبنية على ملاحظة أنه لو تطوى البروتين عبر اتخاذ جميع الهيئات الممكنة تسلسليا (واحدة تلو الأخرى) لاستغرق فعل ذلك كمية فلكية من الزمن، حتى لو كان الوقت بين اتخاذ هيئتين سريعا جدا (نانو ثانية أو بيكو ثانية).[70] بناء على ملاحظة أن البروتينات تتطوى بسرعة أكبر من ذلك، اقترح ليفينثال أنه لا يحدث بحث عشوائي عن هيئة معينة وأن البروتين نتيجة لذلك يتطوى عبر سلسلة من الحالات الوسيطة شبه المستقرة.
مشهد الطاقة العام لتطوي البروتين
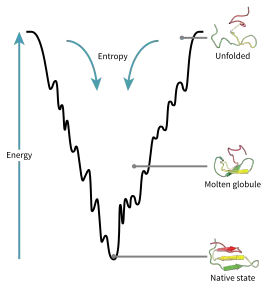
يُصوَّر تشكيل الفضاء [الإنجليزية] الخاص ببروتين أثناء التطوي كمشهد عام للطاقة. تبعا لجوزيف برينجلسون وبيتر وولينز، تتبع البروتينات مبدأ الإحباط الأدنى ويعني ذلك أن البروتينات المتطورة طبيعيا تم إمثال (جعلها الأمثل) مشاهد الطاقة العامة للتطوي الخاصة بها،[71] وأن الطبيعة اختارت تسلسل أحماض أمينية بحيث تكون الهيئة المتطوية من البروتين مستقرة كفاية، وأيضا عملية اتخاذ هيئة التطوي توجب عليها أن تكون عملية سريعة كفاية كذلك. رغم أن الطبيعة خفضت من مستوى الإحباط في البروتينات، فإن نسبة منه مازالت باقية لحد الآن، ويمكن ملاحظة ذلك في تواجد النقطة المحلية الصغرى في مشهد الطاقة العام للبروتينات.
من نتائج هذه التسلسلات المصطفاة تطوريا الاعتقاد بأن للبروتينات «مشهد طاقة عام على شكل قمع» (من صياغة جوسيه أونوتشيك)[72] موجه بشكل كبير نحو الهيئة الطبيعية. يسمح المشهد العام الخاص "بقُمع التطوي [الإنجليزية]" للبروتين بالتطوي إلى الهيئة الطبيعية عبر أي مسارٍ وهيئةٍ وسطيةٍ من بين أعداد كبيرة من المسارات والهيئات الوسطية، بدل أن يكون محصورا في آلية واحدة. هذه الفرضية مدعومة من قبل المحاكيات الحاسوية لنموذج البروتينات [الإنجليزية] والدراسات التجريبية،[71] وتُستخدم لتحسين طرق التنبؤ ببنية البروتين وتصميمه.[71] وصف تطوي البروتين بواسطة تحديد مستوى الطاقة الحرة في مشهد الطاقة العم متسق كذلك مع القانون الثاني للديناميكا الحرارية.[73] فيزيائيا، التفكير في المشاهد العامة من منطلق أنها أسطح طاقة كاملة أو كامنة تملك نقاطا عظمى ودنيا وسرجية -تماما مثل المناظر العامة الجغرافية- أمر مضلل قليلا ربما. الوصف المناسب في الحقيقة هو فضاء طور متعدد الأبعاد قد تأخذ فيه المتنوعات هيئات طوبولوجية متنوعة أكثر تعقيدا.[74]
تبدأ سلاسل عديد الببتيد غير المتطوية في قمة القمع أين يمكنها اتخاذ عدد كبير من الهيئات غير المتطوية وتكون في أقصى حالتها الطاقوية. تشير المشاهد العامة للطاقة إلى وجود عدد كبير من الهيئات الممكنة في البداية وعدم وجود سوى هيئة طبيعية (واطنة) واحدة، ومع ذلك لا تكشف مسارات التطوي المتعددة الممكنة. يمكن أن يتبع جزيئان من نفس البروتين مسارَيْ تطوي مختلفين قليلا، ويسعيان نحو هيئات وسطية مختلفة أقل طاقة، طالما يصلان في النهاية إلى نفس البنية الطبيعية.[75] قد تُستخدم مسارات معينة بشكل أكبر من مسارات أخرى وذلك حسب التفضيل الثرموديناميكي لكل مسار، وهذا يعني أن المسار الذي يكون مفضلا ثرموديناميكيا عن الآخر فعلى الأرجح سيتم استخدامه بتواتر أكبر في سبيل الوصول للبنية الطبيعية.[75] بينما يبدأ البروتين في التطوي واتخاذ عدة هيئات وسطية، يسعى دائما نحو البنية الأكثر تفضيلا ثرموديناميكيا من البنية التي سبقتها ويستمر كذلك عبر قمع الطاقة. تَكوُّن البنية الثانوية مؤشرٌ قوي على زيادة الاستقرار ذاخل البروتين، ولا يمكن سوى لتوليفة واحدة من البُنى الثانوية التي يتطوي إليها عمود عديد الببتيد الفقري أن تكون لديها حالة طاقوية أخفض وبالتالي تكون موجودة أثناء الوصول إلى هيئة التطوي الطبيعية.[75] من بين البنى الأولى التي تتشكل عندما يبدأ عديد الببتيد في التطوي هي: لوالب ألفا التي تتشكل في حوالي 100 نانو ثانية ولفات بيتا [الإنجليزية] التي تتشكل في 1 ميلي ثانية.[28]
توجد نقطة سرجية في المنظر العام لقُمع الطاقة وفيها تتواجد حالة انتقالية لبروتين معين.[28] الحالة الانتقالية في مخطط قمع الطاقة هي الهيئة التي يجب أن يتخذها كل جزيء من ذلك البروتين إذا أراد البروتين أن يصل إلى بنيته الطبيعية في النهاية. لا يمكن لأي بروتين اتخاذ البنية الطبيعية مباشرة دون المرور عبر حالة انتقالية.[28] يمكن الإشارة إلى الحالة الانتقالية على أنها هيئة مغايرة أو مبتسرة من الهيئة الطبيعية بدل اعتبارها مجرد خطوة وسطية أخرى نحوها.[76] تم إظهار أن تطوي الحالة الانتقالية محدِّدٌ لمعدل التطوي -ورغم أنها تتواجد في حالة طاقوية أعلى من الهيئة الطبيعية- فإنها تماثل الهيئة الطبيعية كثيرا. يتواجد داخل الحالة الانتقالية نواةٌ يمكن للبروتين التطوي حولها وتتشكل هذه النواة عبر عملية تسمى «تكثف التنوي» أين تبدأ فيها البنية بالانهيار نحو النواة.[76]
نمذجة تطوي البروتين
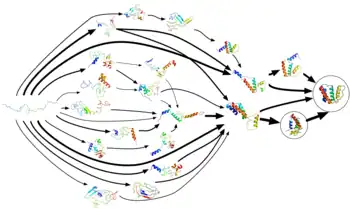
يمكن أن تُستخدم تقنيات الحوسبة الجديدة والبدئية الخاصة بالتنبؤ ببنية البروتين لمحاكاة مختلف نواحي تطوي البروتين. استُخدمت الديناميكا الجزيئية في محاكَيَاتٍ (جمع محاكاة) لتطوي البروتين وحركياته حاسوبيا.[77] نُفذت أول محاكَيات لتوازن التطوي باستخدام نموذج المذيب واعتيان المظلة [الإنجليزية].[78] بسبب تكلفة الحوسبة، محاكيات الديناميكا الجزيئية البدئية مع ماء [الإنجليزية] صريح محدودة في الببتيدات والبروتينات الصغيرة.[79][80] تبقى محاكيات الديناميكا الجزيئية للبروتينات الكبيرة محصورة في حركيات البنية التجريبية أو فك تطويها بسبب ارتفاع الحرارة. يمكن الوصول لعمليات التطوي طويلة الأمد (أكبر من 1 ميلي ثانية) مثل تطوي البروتينات صغيرة الحجم (حوالي خمسين حمض أميني) أو أكبر عبر استخدام نماذج التحبيب التقريبي [الإنجليزية].[81][82][83]
تستهدف عدة مشاريع حوسبة كبيرة مثل: روستا آت هوم،[84] فولدنغ@هوم،[85] وفولديت [الإنجليزية][86] مسألة تطوي البروتين.
تم تنفيذ محاكيات المسار الطويلة المستمرة على حاسوب أنتون [الإنجليزية]، وهو حاسوب فائق متوازي ضخم، تم بناؤه من دارات مدمجة مخصصة التطبيق [الإنجليزية] (ASICs) وروابط [الإنجليزية] بواسطة شركة (D. E. Shaw Research). أكبر نتيجة منشورة لمحاكاة باستخدام أنتون هي 2.936 ميلي ثانية لبروتين NTL9.[87]
انظر أيضًا
المراجع
- Alberts B، Johnson A، Lewis J، Raff M، Roberts K، Walters P (2002). "The Shape and Structure of Proteins". Molecular Biology of the Cell; Fourth Edition. New York and London: Garland Science. ISBN:978-0-8153-3218-3.
- Anfinsen CB (يوليو 1972). "The formation and stabilization of protein structure". The Biochemical Journal. ج. 128 ع. 4: 737–49. DOI:10.1042/bj1280737. PMC:1173893. PMID:4565129.
- Berg JM، Tymoczko JL، Stryer L (2002). "3. Protein Structure and Function". Biochemistry. San Francisco: W. H. Freeman. ISBN:978-0-7167-4684-3.
- Selkoe DJ (ديسمبر 2003). "Folding proteins in fatal ways". Nature. ج. 426 ع. 6968: 900–4. Bibcode:2003Natur.426..900S. DOI:10.1038/nature02264. PMID:14685251. S2CID:6451881.
- Alberts B، Bray D، Hopkin K، Johnson A، Lewis J، Raff M، Roberts K، Walter P (2010). "Protein Structure and Function". Essential cell biology (ط. Third). New York, NY: Garland Science. ص. 120–70. ISBN:978-0-8153-4454-4.
- Kim PS، Baldwin RL (1990). "Intermediates in the folding reactions of small proteins". Annual Review of Biochemistry. ج. 59: 631–60. DOI:10.1146/annurev.bi.59.070190.003215. PMID:2197986.
- Jackson SE (1998). "How do small single-domain proteins fold?". Folding & Design. ج. 3 ع. 4: R81-91. DOI:10.1016/S1359-0278(98)00033-9. PMID:9710577.
- Kubelka J، Hofrichter J، Eaton WA (فبراير 2004). "The protein folding 'speed limit'". Current Opinion in Structural Biology. ج. 14 ع. 1: 76–88. DOI:10.1016/j.sbi.2004.01.013. PMID:15102453. مؤرشف من الأصل في 2020-08-06.
- Anfinsen CB (يوليو 1973). "Principles that govern the folding of protein chains". Science. ج. 181 ع. 4096: 223–30. Bibcode:1973Sci...181..223A. DOI:10.1126/science.181.4096.223. PMID:4124164.
- Voet D، Voet JG، Pratt CW (2016). Principles of Biochemistry (ط. Fifth). Wiley. ISBN:978-1-118-91840-1.
- Alexander PA، He Y، Chen Y، Orban J، Bryan PN (يوليو 2007). "The design and characterization of two proteins with 88% sequence identity but different structure and function". Proceedings of the National Academy of Sciences of the United States of America. ج. 104 ع. 29: 11963–8. Bibcode:2007PNAS..10411963A. DOI:10.1073/pnas.0700922104. PMC:1906725. PMID:17609385.
- Rose GD، Fleming PJ، Banavar JR، Maritan A (نوفمبر 2006). "A backbone-based theory of protein folding". Proceedings of the National Academy of Sciences of the United States of America. ج. 103 ع. 45: 16623–33. Bibcode:2006PNAS..10316623R. CiteSeerX:10.1.1.630.5487. DOI:10.1073/pnas.0606843103. PMC:1636505. PMID:17075053.
- Fersht A (1999). Structure and Mechanism in Protein Science: A Guide to Enzyme Catalysis and Protein Folding. Macmillan. ISBN:978-0-7167-3268-6. مؤرشف من الأصل في 2021-01-04.
- "Protein Structure". Scitable. Nature Education. مؤرشف من الأصل في 2020-10-08. اطلع عليه بتاريخ 2016-11-26.
- Pratt C، Cornely K (2004). "Thermodynamics". Essential Biochemistry. Wiley. ISBN:978-0-471-39387-0. مؤرشف من الأصل في 2020-11-08. اطلع عليه بتاريخ 2016-11-26.
- Zhang G، Ignatova Z (فبراير 2011). "Folding at the birth of the nascent chain: coordinating translation with co-translational folding". Current Opinion in Structural Biology. ج. 21 ع. 1: 25–31. DOI:10.1016/j.sbi.2010.10.008. PMID:21111607.
- van den Berg B، Wain R، Dobson CM، Ellis RJ (أغسطس 2000). "Macromolecular crowding perturbs protein refolding kinetics: implications for folding inside the cell". The EMBO Journal. ج. 19 ع. 15: 3870–5. DOI:10.1093/emboj/19.15.3870. PMC:306593. PMID:10921869.
- Al-Karadaghi S. "Torsion Angles and the Ramachnadran Plot in Protein Structures". www.proteinstructures.com. مؤرشف من الأصل في 2020-08-06. اطلع عليه بتاريخ 2016-11-26.
- Pace CN، Shirley BA، McNutt M، Gajiwala K (يناير 1996). "Forces contributing to the conformational stability of proteins". FASEB Journal. ج. 10 ع. 1: 75–83. DOI:10.1096/fasebj.10.1.8566551. PMID:8566551. S2CID:20021399.
- Cui D، Ou S، Patel S (ديسمبر 2014). "Protein-spanning water networks and implications for prediction of protein-protein interactions mediated through hydrophobic effects". Proteins. ج. 82 ع. 12: 3312–26. DOI:10.1002/prot.24683. PMID:25204743. S2CID:27113763.
- Tanford C (يونيو 1978). "The hydrophobic effect and the organization of living matter". Science. ج. 200 ع. 4345: 1012–8. Bibcode:1978Sci...200.1012T. DOI:10.1126/science.653353. PMID:653353.
- Deechongkit S، Nguyen H، Powers ET، Dawson PE، Gruebele M، Kelly JW (يوليو 2004). "Context-dependent contributions of backbone hydrogen bonding to beta-sheet folding energetics". Nature. ج. 430 ع. 6995: 101–5. Bibcode:2004Natur.430..101D. DOI:10.1038/nature02611. PMID:15229605. S2CID:4315026.
- Irbäck A، Sandelin E (نوفمبر 2000). "On hydrophobicity correlations in protein chains". Biophysical Journal. ج. 79 ع. 5: 2252–8. arXiv:cond-mat/0010390. Bibcode:2000BpJ....79.2252I. DOI:10.1016/S0006-3495(00)76472-1. PMC:1301114. PMID:11053106.
- Irbäck A، Peterson C، Potthast F (سبتمبر 1996). "Evidence for nonrandom hydrophobicity structures in protein chains". Proceedings of the National Academy of Sciences of the United States of America. ج. 93 ع. 18: 9533–8. arXiv:chem-ph/9512004. Bibcode:1996PNAS...93.9533I. DOI:10.1073/pnas.93.18.9533. PMC:38463. PMID:8790365.
- Wilson BA، Foy SG، Neme R، Masel J (يونيو 2017). "De Novo Gene Birth". Nature Ecology & Evolution. ج. 1 ع. 6: 0146–146. DOI:10.1038/s41559-017-0146. PMC:5476217. PMID:28642936.
- Willis S، Masel J (سبتمبر 2018). "Gene Birth Contributes to Structural Disorder Encoded by Overlapping Genes". Genetics. ج. 210 ع. 1: 303–313. DOI:10.1534/genetics.118.301249. PMC:6116962. PMID:30026186.
- Foy SG، Wilson BA، Bertram J، Cordes MH، Masel J (أبريل 2019). "A Shift in Aggregation Avoidance Strategy Marks a Long-Term Direction to Protein Evolution". Genetics. ج. 211 ع. 4: 1345–1355. DOI:10.1534/genetics.118.301719. PMC:6456324. PMID:30692195.
- Dobson CM (ديسمبر 2003). "Protein folding and misfolding". Nature. ج. 426 ع. 6968: 884–90. Bibcode:2003Natur.426..884D. DOI:10.1038/nature02261. PMID:14685248. S2CID:1036192.
- Hartl FU (يونيو 1996). "Molecular chaperones in cellular protein folding". Nature. ج. 381 ع. 6583: 571–9. Bibcode:1996Natur.381..571H. DOI:10.1038/381571a0. PMID:8637592. S2CID:4347271.
- Hartl FU، Bracher A، Hayer-Hartl M (يوليو 2011). "Molecular chaperones in protein folding and proteostasis". Nature. ج. 475 ع. 7356: 324–32. DOI:10.1038/nature10317. PMID:21776078. S2CID:4337671.
- Kim YE، Hipp MS، Bracher A، Hayer-Hartl M، Hartl FU (2013). "Molecular chaperone functions in protein folding and proteostasis". Annual Review of Biochemistry. ج. 82: 323–55. DOI:10.1146/annurev-biochem-060208-092442. PMID:23746257.
- Shortle D (يناير 1996). "The denatured state (the other half of the folding equation) and its role in protein stability". FASEB Journal. ج. 10 ع. 1: 27–34. DOI:10.1096/fasebj.10.1.8566543. PMID:8566543. S2CID:24066207.
- Lee S، Tsai FT (2005). "Molecular chaperones in protein quality control". Journal of Biochemistry and Molecular Biology. ج. 38 ع. 3: 259–65. DOI:10.5483/BMBRep.2005.38.3.259. PMID:15943899.
- Ojeda-May P، Garcia ME (يوليو 2010). "Electric field-driven disruption of a native beta-sheet protein conformation and generation of a helix-structure". Biophysical Journal. ج. 99 ع. 2: 595–9. Bibcode:2010BpJ....99..595O. DOI:10.1016/j.bpj.2010.04.040. PMC:2905109. PMID:20643079.
- van den Berg B، Ellis RJ، Dobson CM (ديسمبر 1999). "Effects of macromolecular crowding on protein folding and aggregation". The EMBO Journal. ج. 18 ع. 24: 6927–33. DOI:10.1093/emboj/18.24.6927. PMC:1171756. PMID:10601015.
- Ellis RJ (يوليو 2006). "Molecular chaperones: assisting assembly in addition to folding". Trends in Biochemical Sciences. ج. 31 ع. 7: 395–401. DOI:10.1016/j.tibs.2006.05.001. PMID:16716593.
- Takai K، Nakamura K، Toki T، Tsunogai U، Miyazaki M، Miyazaki J، Hirayama H، Nakagawa S، Nunoura T، Horikoshi K (أغسطس 2008). "Cell proliferation at 122 degrees C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation". Proceedings of the National Academy of Sciences of the United States of America. ج. 105 ع. 31: 10949–54. Bibcode:2008PNAS..10510949T. DOI:10.1073/pnas.0712334105. PMC:2490668. PMID:18664583.
- Marusich EI, Kurochkina LP, Mesyanzhinov VV. Chaperones in bacteriophage T4 assembly. Biochemistry (Mosc). 1998;63(4):399-406
- Porter، Lauren L.؛ Looger، Loren L. (5 يونيو 2018). "Extant fold-switching proteins are widespread". Proceedings of the National Academy of Sciences. ج. 115 ع. 23: 5968–5973. DOI:10.1073/pnas.1800168115.
- Chaudhuri TK، Paul S (أبريل 2006). "Protein-misfolding diseases and chaperone-based therapeutic approaches". The FEBS Journal. ج. 273 ع. 7: 1331–49. DOI:10.1111/j.1742-4658.2006.05181.x. PMID:16689923. S2CID:23370420.
- Maji، Samir K.؛ Wang، Lei؛ Greenwald، Jason؛ Riek، Roland (11 يوليو 2009). "Structure-activity relationship of amyloid fibrils". FEBS Letters. Wiley. ج. 583 ع. 16: 2610–2617. DOI:10.1016/j.febslet.2009.07.003. ISSN:0014-5793.
- Soto C، Estrada L، Castilla J (مارس 2006). "Amyloids, prions and the inherent infectious nature of misfolded protein aggregates". Trends in Biochemical Sciences. ج. 31 ع. 3: 150–5. DOI:10.1016/j.tibs.2006.01.002. PMID:16473510.
- Hammarström P، Wiseman RL، Powers ET، Kelly JW (يناير 2003). "Prevention of transthyretin amyloid disease by changing protein misfolding energetics". Science. ج. 299 ع. 5607: 713–6. Bibcode:2003Sci...299..713H. DOI:10.1126/science.1079589. PMID:12560553. S2CID:30829998.
- Chiti F، Dobson CM (2006). "Protein misfolding, functional amyloid, and human disease". Annual Review of Biochemistry. ج. 75: 333–66. DOI:10.1146/annurev.biochem.75.101304.123901. PMID:16756495.
- Johnson SM، Wiseman RL، Sekijima Y، Green NS، Adamski-Werner SL، Kelly JW (ديسمبر 2005). "Native state kinetic stabilization as a strategy to ameliorate protein misfolding diseases: a focus on the transthyretin amyloidoses". Accounts of Chemical Research. ج. 38 ع. 12: 911–21. DOI:10.1021/ar020073i. PMID:16359163.
- Cowtan K (2001). "Phase Problem in X-ray Crystallography, and Its Solution" (PDF). Encyclopedia of Life Sciences. Macmillan Publishers Ltd, Nature Publishing Group. مؤرشف من الأصل (PDF) في 2020-08-06. اطلع عليه بتاريخ 2016-11-03.
- Drenth J (5 أبريل 2007). Principles of Protein X-Ray Crystallography. Springer Science & Business Media. ISBN:978-0-387-33746-3. مؤرشف من الأصل في 2021-01-04.
- Taylor G (2003). "The phase problem". Acta Crystallographica Section D. ج. 59 ع. 11: 1881–90. DOI:10.1107/S0907444903017815. PMID:14573942.
- Bedouelle H (فبراير 2016). "Principles and equations for measuring and interpreting protein stability: From monomer to tetramer". Biochimie. ج. 121: 29–37. DOI:10.1016/j.biochi.2015.11.013. PMID:26607240.
- Monsellier E، Bedouelle H (سبتمبر 2005). "Quantitative measurement of protein stability from unfolding equilibria monitored with the fluorescence maximum wavelength". Protein Engineering, Design & Selection. ج. 18 ع. 9: 445–56. DOI:10.1093/protein/gzi046. PMID:16087653.
- Park YC، Bedouelle H (يوليو 1998). "Dimeric tyrosyl-tRNA synthetase from Bacillus stearothermophilus unfolds through a monomeric intermediate. A quantitative analysis under equilibrium conditions". The Journal of Biological Chemistry. ج. 273 ع. 29: 18052–9. DOI:10.1074/jbc.273.29.18052. PMID:9660761.
- Ould-Abeih MB، Petit-Topin I، Zidane N، Baron B، Bedouelle H (يونيو 2012). "Multiple folding states and disorder of ribosomal protein SA, a membrane receptor for laminin, anticarcinogens, and pathogens". Biochemistry. ج. 51 ع. 24: 4807–21. DOI:10.1021/bi300335r. PMID:22640394.
- Royer CA (مايو 2006). "Probing protein folding and conformational transitions with fluorescence". Chemical Reviews. ج. 106 ع. 5: 1769–84. DOI:10.1021/cr0404390. PMID:16683754.
- Wüthrich K (ديسمبر 1990). "Protein structure determination in solution by NMR spectroscopy". The Journal of Biological Chemistry. ج. 265 ع. 36: 22059–62. PMID:2266107. مؤرشف من الأصل في 2020-01-17.
- Zhuravleva A، Korzhnev DM (مايو 2017). "Protein folding by NMR". Progress in Nuclear Magnetic Resonance Spectroscopy. ج. 100: 52–77. DOI:10.1016/j.pnmrs.2016.10.002. PMID:28552172. مؤرشف من الأصل في 2021-01-04.
- Ortega G، Pons M، Millet O (1 يناير 2013). "Protein functional dynamics in multiple timescales as studied by NMR spectroscopy". Advances in Protein Chemistry and Structural Biology. Dynamics of Proteins and Nucleic Acids. Academic Press. ج. 92: 219–51. DOI:10.1016/b978-0-12-411636-8.00006-7. ISBN:9780124116368. PMID:23954103.
- Vallurupalli P، Bouvignies G، Kay LE (مايو 2012). "Studying "invisible" excited protein states in slow exchange with a major state conformation". Journal of the American Chemical Society. ج. 134 ع. 19: 8148–61. DOI:10.1021/ja3001419. PMID:22554188.
- Neudecker P، Lundström P، Kay LE (مارس 2009). "Relaxation dispersion NMR spectroscopy as a tool for detailed studies of protein folding". Biophysical Journal. ج. 96 ع. 6: 2045–54. Bibcode:2009BpJ....96.2045N. DOI:10.1016/j.bpj.2008.12.3907. PMC:2717354. PMID:19289032.
- Sekhar A، Rumfeldt JA، Broom HR، Doyle CM، Sobering RE، Meiering EM، Kay LE (نوفمبر 2016). "Probing the free energy landscapes of ALS disease mutants of SOD1 by NMR spectroscopy". Proceedings of the National Academy of Sciences of the United States of America. ج. 113 ع. 45: E6939–E6945. DOI:10.1073/pnas.1611418113. PMC:5111666. PMID:27791136.
- Cross GH، Freeman NJ، Swann MJ (2008). "Dual Polarization Interferometry: A Real-Time Optical Technique for Measuring (Bio)molecular Orientation, Structure and Function at the Solid/Liquid Interface". Handbook of Biosensors and Biochips. DOI:10.1002/9780470061565.hbb055. ISBN:978-0-470-01905-4.
- Bu Z، Cook J، Callaway DJ (سبتمبر 2001). "Dynamic regimes and correlated structural dynamics in native and denatured alpha-lactalbumin". Journal of Molecular Biology. ج. 312 ع. 4: 865–73. DOI:10.1006/jmbi.2001.5006. PMID:11575938.
- Minde DP، Maurice MM، Rüdiger SG (2012). "Determining biophysical protein stability in lysates by a fast proteolysis assay, FASTpp". PLOS ONE. ج. 7 ع. 10: e46147. Bibcode:2012PLoSO...746147M. DOI:10.1371/journal.pone.0046147. PMC:3463568. PMID:23056252.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - Park C، Marqusee S (مارس 2005). "Pulse proteolysis: a simple method for quantitative determination of protein stability and ligand binding". Nature Methods. ج. 2 ع. 3: 207–12. DOI:10.1038/nmeth740. PMID:15782190. S2CID:21364478.
- Mashaghi A، Kramer G، Lamb DC، Mayer MP، Tans SJ (يناير 2014). "Chaperone action at the single-molecule level". Chemical Reviews. ج. 114 ع. 1: 660–76. DOI:10.1021/cr400326k. PMID:24001118.
- Jagannathan B، Marqusee S (نوفمبر 2013). "Protein folding and unfolding under force". Biopolymers. ج. 99 ع. 11: 860–9. DOI:10.1002/bip.22321. PMC:4065244. PMID:23784721.
- Jakobi AJ، Mashaghi A، Tans SJ، Huizinga EG (يوليو 2011). "Calcium modulates force sensing by the von Willebrand factor A2 domain". Nature Communications. ج. 2: 385. Bibcode:2011NatCo...2..385J. DOI:10.1038/ncomms1385. PMC:3144584. PMID:21750539.
- Jagannathan B، Elms PJ، Bustamante C، Marqusee S (أكتوبر 2012). "Direct observation of a force-induced switch in the anisotropic mechanical unfolding pathway of a protein". Proceedings of the National Academy of Sciences of the United States of America. ج. 109 ع. 44: 17820–5. Bibcode:2012PNAS..10917820J. DOI:10.1073/pnas.1201800109. PMC:3497811. PMID:22949695.
- Compiani M، Capriotti E (ديسمبر 2013). "Computational and theoretical methods for protein folding". Biochemistry. ج. 52 ع. 48: 8601–24. DOI:10.1021/bi4001529. PMID:24187909.
- "Structural Biochemistry/Proteins/Protein Folding - Wikibooks, open books for an open world". en.wikibooks.org. مؤرشف من الأصل في 2020-10-08. اطلع عليه بتاريخ 2016-11-05.
- Levinthal C (1968). "Are there pathways for protein folding?" (PDF). Journal de Chimie Physique et de Physico-Chimie Biologique. ج. 65: 44–45. Bibcode:1968JCP....65...44L. DOI:10.1051/jcp/1968650044. مؤرشف من الأصل (PDF) في 2009-09-02.
- Bryngelson JD، Onuchic JN، Socci ND، Wolynes PG (مارس 1995). "Funnels, pathways, and the energy landscape of protein folding: a synthesis". Proteins. ج. 21 ع. 3: 167–95. arXiv:chem-ph/9411008. DOI:10.1002/prot.340210302. PMID:7784423. S2CID:13838095.
- Leopold PE، Montal M، Onuchic JN (سبتمبر 1992). "Protein folding funnels: a kinetic approach to the sequence-structure relationship". Proceedings of the National Academy of Sciences of the United States of America. ج. 89 ع. 18: 8721–5. Bibcode:1992PNAS...89.8721L. DOI:10.1073/pnas.89.18.8721. PMC:49992. PMID:1528885.
- Sharma V، Kaila VR، Annila A (2009). "Protein folding as an evolutionary process". Physica A: Statistical Mechanics and Its Applications. ج. 388 ع. 6: 851–62. Bibcode:2009PhyA..388..851S. DOI:10.1016/j.physa.2008.12.004.
- Robson B، Vaithilingam A (2008). "Protein Folding Revisited". Molecular Biology of Protein Folding, Part B. Progress in Molecular Biology and Translational Science. ج. 84. ص. 161–202. DOI:10.1016/S0079-6603(08)00405-4. ISBN:978-0-12-374595-8. PMID:19121702.
- Dill KA، MacCallum JL (نوفمبر 2012). "The protein-folding problem, 50 years on". Science. ج. 338 ع. 6110: 1042–6. Bibcode:2012Sci...338.1042D. DOI:10.1126/science.1219021. PMID:23180855. S2CID:5756068.
- Fersht AR (فبراير 2000). "Transition-state structure as a unifying basis in protein-folding mechanisms: contact order, chain topology, stability, and the extended nucleus mechanism". Proceedings of the National Academy of Sciences of the United States of America. ج. 97 ع. 4: 1525–9. Bibcode:2000PNAS...97.1525F. DOI:10.1073/pnas.97.4.1525. PMC:26468. PMID:10677494.
- Rizzuti B، Daggett V (مارس 2013). "Using simulations to provide the framework for experimental protein folding studies". Archives of Biochemistry and Biophysics. ج. 531 ع. 1–2: 128–35. DOI:10.1016/j.abb.2012.12.015. PMC:4084838. PMID:23266569.
- Schaefer M، Bartels C، Karplus M (ديسمبر 1998). "Solution conformations and thermodynamics of structured peptides: molecular dynamics simulation with an implicit solvation model". Journal of Molecular Biology. ج. 284 ع. 3: 835–48. DOI:10.1006/jmbi.1998.2172. PMID:9826519.
- Jones D. "Fragment-based Protein Folding Simulations". University College London. مؤرشف من الأصل في 2020-08-09.
- "Protein folding". مؤرشف من الأصل (by Molecular Dynamics) في 2020-07-05.
- Kmiecik S، Gront D، Kolinski M، Wieteska L، Dawid AE، Kolinski A (يوليو 2016). "Coarse-Grained Protein Models and Their Applications". Chemical Reviews. ج. 116 ع. 14: 7898–936. DOI:10.1021/acs.chemrev.6b00163. PMID:27333362.
- Kmiecik S، Kolinski A (يوليو 2007). "Characterization of protein-folding pathways by reduced-space modeling". Proceedings of the National Academy of Sciences of the United States of America. ج. 104 ع. 30: 12330–5. Bibcode:2007PNAS..10412330K. DOI:10.1073/pnas.0702265104. PMC:1941469. PMID:17636132.
- Adhikari AN، Freed KF، Sosnick TR (أكتوبر 2012). "De novo prediction of protein folding pathways and structure using the principle of sequential stabilization". Proceedings of the National Academy of Sciences of the United States of America. ج. 109 ع. 43: 17442–7. Bibcode:2012PNAS..10917442A. DOI:10.1073/pnas.1209000109. PMC:3491489. PMID:23045636.
- Rosetta@Home نسخة محفوظة 4 يناير 2021 على موقع واي باك مشين.
- Folding@Home نسخة محفوظة 21 نوفمبر 2020 على موقع واي باك مشين.
- FoldIt - Folding Protein Game نسخة محفوظة 4 يناير 2021 على موقع واي باك مشين.
- Lindorff-Larsen K، Piana S، Dror RO، Shaw DE (أكتوبر 2011). "How fast-folding proteins fold". Science. ج. 334 ع. 6055: 517–20. Bibcode:2011Sci...334..517L. DOI:10.1126/science.1208351. PMID:22034434. S2CID:27988268.
 بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي