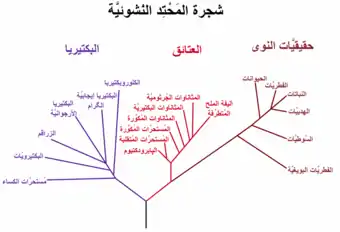
شجرة تطور السلالات
شجرة المَحْتِد (بالإنجليزية: phylogenetic tree) أو شجرة التطور هي عبارة عن شجرة تظهر العلاقات التطورية بين مختلف الأنواع الحيوية أو مختلف الكيانات الحية التي يعتقد بأنها تمتلك أصلاً مشتركاً.[1][2][3] تمثل كل عقدة مع تفرعاتها في شجرة المحتد السلف المشترك الأحدث most recent common ancestor لما يتفرع عنه، وأطوال الفروع تمثل تقديرات زمنية. كل عقدة تدعى وحدة أصنوفية taxonomic unit. العقد الداخلية تدعى عادة عقد أصنوفية افتراضية (HTUs) لأنها لا يمكن ملاحظتها بشكل مباشر.
التاريخ
نشأت فكرة "شجرة الحياة" من المفاهيم القديمة للتعاقب السلّمي للكائنات الحيّة من الأشكال السفلى للحياة إلى الأشكال العليا (كما في سلسلة الكينونة العظمى، أو سلسلة الوجود). تتضمن التصوّرات المبكّرة لشجرة الحياة «المتفرّعة» أيضاً «مخططًا قديمًا» يوضّح العلاقات الجيولوجية بين النباتات والحيوانات تمّ وضعه من قبل إدوارد هيتشكوك في كتابه الجيولوجيا البسيطة (الذي نشر في طبعته الأولى: 1840).
قام تشارلز داروين (1859) أيضًا بوضع واحد من الرسوم التوضيحية الأولى ودَعَمَ مفهوم «شجرة التطوّر» وذلك في كتابه «أصل الأنواع». لا يزال علماء الأحياء التطوّرية -بعد أكثر من قرن من الزمان على نشر الكتاب- يستخدمون المخطّطات الشجريّة لتصوير التطوّر، لأنّها تنقل المفهوم القائل بأنّ التكاثر يحدث من خلال التقسيم التكيفي وشبه العشوائي للسلالات بفعّاليّة عالية. أصبح تصنيف الأنواع أقل جموداً وأكثر ديناميكية مع مرور الزمن.
مصطلح phylogenetic «متعلق بتَطَوُّرِ السُّلاَلاَت»، أو phylogeny «علمُ تَطَوُّرِ السُّلاَلاَت»، مشتق من الكلمتين اليونانيتين القديمتين φῦλονالتي تلفظ (phûlon)، بمعنى «العرق، والنسب»، و γένεσις التي تلفظ (génesis)، بمعنى «الأصل، المصدر».[4][5]
الأنواع
الشجرة ذات الجذر (الأصل) المشترك
الشجرة ذات الأصل المشترك (انظر الرسومات في الأعلى) هي شجرة موجّهة ذات نقطة التقاء فريدة -الجذر- والتي تقابل السلف المشترك الأوّل الذي (عادةً ما يتم اعتباره) الأصل المشترك الأحدث لجميع الكيانات الحيّة الموجودة على فروع وأوراق الشجرة التطوريّة. لا تملك عقدة الأصل المشترك عقدة أصلية، ولكنها بمثابة الأصل لجميع العقد الأخرى في الشجرة التطوّريّة. وبالتالي فإنّ الجذر هو عقدة من الدرجة 2 بينما العقد الداخلية الأخرى لديها درجة لا تقل عن 3 (حيث تشير «درجة» هنا إلى العدد الإجمالي للخطوط الواردة والصادرة للعقدة).
تتمثّل الطريقة الأكثر شيوعًا لوضع الجذر (الأصل) للشجرة التطوّريّة باستخدام مجموعة خارجية غير مثيرة للجدل بمعنى أن تكون قريبة بدرجة كافية للسماح بالاستدلال عن طريق بيانات السمات أو التسلسل الجزيئي، ولكن بالمقابل بعيدة بدرجة كافية لتكون مجموعة خارجية واضحة.
شجرة التطور بلا جذر (أصل) مشترك
توضّح الأشجار التي لا تحوي على جذر (أصل) مشترك العلاقة بين العقد دون وضع افتراضات حول نسبها. فهذا النوع من أشجار التطور لا يتطلّب أن يكون الأصل معروفًا أو مستنتجًا.[6] يمكن دائمًا أن تنتج الأشجار التي لا تحوي أصلاً مشتركاً عن الأشجار ذات الأصل المشترك بحذف الأصل. لكن، وعلى النقيض من ذلك، فإنّ استنباط الأصل لشجرة لا تملك أصلاً مشتركاً يتطلّب بعض الوسائل لتحديده. ويتمّ ذلك عادةً عن طريق تضمين مجموعة خارجيّة في بيانات الإدخال وبذلك يكون الأصل بين المجموعة الخارجية وباقي الأصناف الموجودة في الشجرة بالضرورة، أو عن طريق تقديم افتراضات إضافيّة حول المعدّلات النسبية للتطور في كلّ فرع، مثل التطبيق فرضيّة الساعة الجزيئية.[1]
أنواع خاصة من أشجار تطور السلالات
إن الرسم الشجري هو اسم عام للشجرة، سواء أكانت شجرة سلالات تطورية أم لا، وبالتالي هو اسم عامٌ أيضاً للتمثيل البياني لشجرة تطور السلالات. يمثل مخطط النسل نمطاً تفرعياً فحسب. أي أن امتدادات فروعه لا تمثل زمناً أو مقداراً نسبياً من تغير سمة ما، كما أن العقد الداخلية لا تمثل أسلافاً.
أما الـ phylogram، فهي عبارة عن شجرة تطور السلالات التي تملك امتدادات فرعية تتناسب مع مقدار تغير سمة ما.[7] والكرونوغرام عبارة عن شجرة تطور السلالات التي تمثل الزمن بوضوح خلال امتداداتها الفرعية.[8]
يُعد مخطط المغزل (والذي يُسمى غالباً رسم روميوجرام على اسم عالم الحفريات الأمريكي ألفريد رومر) تمثيلاً لتطور ووفرة الأصناف المختلفة عبر الزمن، ولكنه ليس شجرة تطورية. أما Dahlgrenogram فهو رسم بياني يمثل مقطعاً عرضياً من شجرة تطور السلالات.
وهناك شبكة تطور السلالات، والتي لا تعني بالضرورة شجرة تطور، بل هي رسم بياني أكثر عمومية، أو رسم بياني لا حلقي موجه في الشبكات ذات الجذور. وتُستخدم للتغلب على بعض القيود الملازمة للأشجار.
بناء شجرة تطور السلالات
تُنشأ أشجار تطور السلالات المكونة من عدد غير بديهي من تسلسل المدخلات باستخدام أساليب علم تطور السلالات الحسابي. تعتبر أساليب مصفوفة المسافة، مثل الارتباط المتجاور أو طريقة UPGMA، التي تحسب المسافة الجينية من تراصف السلسلة المتعدد أبسط الطرق من ناحية التنفيذ. ولكنها لا تستند على نموذج تطوري. تقوم العديد من أساليب تراصف التسلسل، كبرنامج كلوستال مثلاً، بإنشاء أشجار تطور السلالات باستخدام الخوارزميات الأكثر بساطة (أي تلك المستندة إلى المسافة) لإنشاء هذه الأشجار. وهناك طريقة أخرى بسيطة لحساب شجرة تطور السلالات، والتي تدعى الاقتصاد الأقصى أو Maximum Parismony. ولكنها تشتمل على نموذجٍ ضمني للتطور. تستخدم الأساليب الأكثر تقدماً معيار الأمثلية لتقدير الاحتمال، غالباً عن طريق استدلال بايزي، وتطبق نموذجاً واضحاً للتطور لتقدير شجرة تطور السلالات. إن تحديد شجرة التطور المثلى باستخدام العديد من هذه التقنيات هو مسألة NP صعبة.[9] لذلك تُستخدم طرق الاستثمال والحدس المهني جنباً إلى جنب مع وظائف تصحيح الشجرة لتحديد إن كانت الشجرة جيدة ومناسبة للبيانات.
يمكن تقييم أساليب بناء الأشجار على أساس عدة معايير:[10]
- الكفاءة (كم من الوقت يستغرق حساب الإجابة؟ وكم الذاكرة التي تحتاجها؟)
- الطاقة (هل تستخدم البيانات استخداماً جيداً، أم تُهدر المعلومات؟)
- الاتساق (هل سنحصل على نفس الإجابة مراراً وتكراراً إن حصلنا في كل مرة على بيانات مختلفة لنفس النموذج؟)
- القوة (هل تتعامل بشكل جيد مع الانتهاكات التي تتعارض مع افتراضات النموذج الأساسي؟)
- قابلية الدحض (هل تنبهنا عندما يكون استخدامها غير مجدٍ؟ أي عند انتهاك الافتراضات؟)
حصلت تقنيات بناء أشجار التطور على اهتمام علماء الرياضيات. وبالإمكان أيضاً بناء الأشجار باستخدام نظرية T.
حدود تحليل تطور السلالات
قد توفر الأشجار التطورية، والمنتجة على أساس الجينات المتسلسلة أو البيانات الجينية في أنواع مختلفة، رؤية تطورية. لكن لهذه التحليلات حدود أو قيود مهمة. وهناك أمر بالغ الأهمية، وهو كون هذه الأشجار التطورية المتولدة غير صحيحة بالضرورة –فهي لا تمثل بالضرورة التاريخ التطوري للأصانيف المشمولة بدقة بالغة. وكما هو الحال مع أي نتيجة علمية، فهي قابلة للدحض من خلال إجراء مزيد من الدراسات (على سبيل المثال: جمع بيانات إضافية وتحليل البيانات الموجودة باستخدام أساليب محسنة). وربما تكون البيانات التي نستند إليها مشوشة.[11] فقد يحدث اختلاطٌ في التحليل نتيجة إعادة التركيب الجيني [12] أو نقل الجينات الأفقي[3] أو التهجين بين الأنواع التي لم تكن قريبة على شجرة التطور قبل حدوث التهجين. وقد يحدث هذا الاختلاط بسبب التطور التقاربي أو التسلسل المحفوظ. وهناك أيضاً مشاكلٌ عند إجراء تحليل على نوع واحد من السمات، مثل مورثة أو بروتين واحد، أو عند التحليل المورفولوجي. ويحدث ذلك لأن أشجار تطور السلالات التي يتم بناؤها من مصدر بيانات آخر غير ذي صلة تختلف غالباً عن الأولى. وبالتالي هناك حاجة ماسة إلى أخذ الحيطة والحذر عند استنتاج علاقات تطور السلالات بين الأنواع. وينطبق ذلك بشكل كبير على المواد الوراثية التي تخضع لنقل الجينات الجانبي وتخليق توافقات جينية جديدة، فقد تمتلك الأنماط الفردانية المختلفة تواريخَ مختلفة أيضاً. في هذه الأنواع من التحليل، تكون الشجرة الناتجة عن تحليل نسبي لجين واحد بمثابة إحصاء وتقديرٍ لتاريخ تطور الجينات (أي شجرة الجينات) وليست لتاريخ تطور الأصناف (أي شجرة الأنواع) التي تم أخذ عينات منها. لكن من الناحية المثالية، يجب أن يكون كلاهما قريباً جداً من الآخر. ولهذا السبب، تستخدم دراسات تطور السلالات عموماً مجموعة من الجينات التي تأتي من مصادر جينية مختلفة (على سبيل المثال: من الميتوكوندريا أو البلاستيد مقابل الجينوم النووي)، أو الجينات التي من المتوقع أن تتطور في ظل أنظمة انتقائية مختلفة، وبالتالي فمن غير المرجح أن ينتج تجانس التقويم والشكل (تنادد خاطئ) عن الانتقاء الطبيعي.
اقرأ أيضا
مراجع
- Maher BA (2002). "Uprooting the Tree of Life". The Scientist. ج. 16: 18. مؤرشف من الأصل في 2003-10-02.
- Hodge T, Cope M (1 أكتوبر 2000). "A myosin family tree". J Cell Sci. ج. 113 ع. 19: 3353–4. PMID:10984423. مؤرشف من الأصل في 2007-09-30.
- Woese C (2002). "On the evolution of cells". Proc Natl Acad Sci USA. ج. 99 ع. 13: 8742–7. Bibcode:2002PNAS...99.8742W. DOI:10.1073/pnas.132266999. PMC:124369. PMID:12077305.
- Bailly، Anatole (1 يناير 1981). Abrégé du dictionnaire grec français. Paris: Hachette. ISBN:978-2010035289. OCLC:461974285.
- Bailly، Anatole. "Greek-french dictionary online". www.tabularium.be. مؤرشف من الأصل في 2014-04-21. اطلع عليه بتاريخ 2018-03-02.
- ""Tree" Facts: Rooted versus Unrooted Trees". مؤرشف من الأصل في 2014-04-14. اطلع عليه بتاريخ 2014-05-26.
- Mayr, Ernst (2009)"Cladistic analysis or cladistic classification?". Journal of Zoological Systematics and Evolutionary Research. 12: 94–128. doi:10.1111/j.1439-0469.1974.tb00160.x..
- Fox، Emily. "The dendrogram". coursea. coursea. مؤرشف من الأصل في 2017-09-28. اطلع عليه بتاريخ 2017-09-28.
- Antonio Soares, Ricardo Râbelo, Alexandre Delbem, Optimization based on phylogram analysis, In Expert Systems with Applications, Volume 78, 2017, Pages 32-50, ISSN 0957-4174, https://doi.org/10.1016/j.eswa.2017.02.012. نسخة محفوظة 2020-08-15 على موقع واي باك مشين.
- Santamaria، R.؛ Theron، R. (26 مايو 2009). "Treevolution: visual analysis of phylogenetic trees". Bioinformatics. ج. 25 ع. 15: 1970–1971. DOI:10.1093/bioinformatics/btp333. PMID:19470585.
- Arenas M, Posada D (2010). "The effect of recombination on the reconstruction of ancestral sequences". Genetics. ج. 184 ع. 4: 1133–1139. DOI:10.1534/genetics.109.113423. PMC:2865913. PMID:20124027.
- Townsend JP, Su Z, Tekle Y (2012). "Phylogenetic Signal and Noise: Predicting the Power of a Data Set to Resolve Phylogeny". Genetics. ج. 61 ع. 5: 835–849. DOI:10.1093/sysbio/sys036. PMID:22389443.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: أسماء متعددة: قائمة المؤلفين (link)
 بوابة علم الأحياء
بوابة علم الأحياء بوابة علم النبات
بوابة علم النبات بوابة تاريخ العلوم
بوابة تاريخ العلوم بوابة علم الأحياء التطوري
بوابة علم الأحياء التطوري