سينثيز بيتا-كيتواسيل- أيه سي بي III
في علم الإنزيمات ، سينثيز بيتا-كيتواسيل -إيه سي بي III ( ر.ت.إ 2.3.1.180
) هو إنزيم يحفز التفاعل الكيميائي الحيوي في البكتيريا والحيوان والإنسان. يشترك في تخليق الأحماض الدهنية. يسمى أيضا FabH.
- أسيتيل-كو-أ + مالونيل-[ بروتين حامل الأسيل ] أسيتو أسيتيل-[ بروتين حامل الأسيل ] + CoA + CO 2

| β-ketoacyl-(acyl-carrier-protein) synthase III | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| أرقام التعريف | |||||||||||
| رقم التصنيف الإنزيمي | 2.3.1.180 | ||||||||||
| رقم التسجيل CAS | 9077-10-5 | ||||||||||
| قواعد البيانات | |||||||||||
| قاعدة بيانات الإنزيم | راجع IntEnz | ||||||||||
| قاعدة بيانات براونشفايغ | راجع BRENDA | ||||||||||
| إكسباسي | راجع NiceZyme | ||||||||||
| موسوعة كيوتو | راجع KEGG | ||||||||||
| ميتاسيك | المسار الأيضي | ||||||||||
| بريام | ملف التعريف | ||||||||||
| تركيب بنك بيانات البروتين | RCSB PDB PDBe PDBsum | ||||||||||
| الأونتولوجيا الجينية | AmiGO / EGO | ||||||||||
| |||||||||||
| 3-Oxoacyl-[acyl-carrier-protein (ACP)] synthase III | |
|---|---|
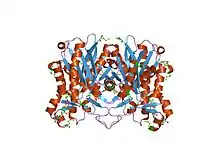 | |
| Structure and active-site architecture of beta-ketoacyl-acyl carrier protein synthase III (FabH) from escherichia coli.[1] | |
| معرف | |
| رمز | ? |
| 3-Oxoacyl- بروتين حامل الأسيل (ACP)] سينسيز III | |
|---|---|
| </img> هيكل وهندسة الموقع النشط لبروتين حامل بروتين بيتا كيتواسيل أسيل سينثاز الثالث (FabH) من الإشريكية القولونية .[2] | |
| معرفات | |
| رمز | ACP_syn_III |
| بفام | PF08545 |
| إنتربرو | IPR013751 |
CoA هو مرافق الإنزيم-أ ويختصر أحيانا في العربيةكو-أ .
وبالتالي ، فإن ركائز هذا الإنزيم هما أسيتيل-كو-أ و مالونيل - [بروتين حامل الأسيل] ، في حين أن منتجاته الثلاثة هي أسيتوأسيتيل-[بروتين حامل الأسيل] و CoA وCO2 . ينتمي هذا الإنزيم إلى عائلة الترانسفيرازات ، ليكون محددًا لتلك المجموعات التي تنقل الأسيلترانسفيرازات غير مجموعات الأمينوأسيل.
يشارك هذا الإنزيم في التخليق الحيوي للأحماض الدهنية . بروتين بيتا-كيتوأسيل-[بروتين حامل الأسيل] III يشارك في نظام التخليق الحيوي للأحماض الدهنية المنفصلة (أو النوع II) الذي يحدث في النباتات والبكتيريا. تم وصف دور FabH في تخليق الأحماض الدهنية في ،Streptomyces glaucescens [3] Streptococcus pneumoniae ، [4] و Streptomyces coelicolor .[5]
التسمية
الاسم المنهجي لفئة الإنزيم هذه هو acetyl-CoA: malonyl- [acyl-carrier-protein] C-acyltransferase. تشمل الأسماء الأخرى الشائعة الاستخدام ما يلي:
|
|
دوره في مرض السل
تتجنب المتفطرة السلية ، التي تسبب مرض السل ، فعل المناعة الفعالة في الإنسان والفطريات من خلال التغليف ، خاصة مع الأحماض الفطرية المقاومة بشكل خاص لعمليات التحلل الطبيعي للبلاعم macrophages . علاوة على ذلك ، تمنع هذه الكبسولة دخول المضادات الحيوية. تعتبر الإنزيمات المشاركة في التخليق الحيوي للميكولات ضرورية للبقاء على قيد الحياة ومحاربة المرض ، وبالتالي تمثل أهدافًا دوائية ممتازة.
في مرض السل ، تم تعيين إنزيم سينثيز بيتا- كيتواسيل-[بروتني حامل الأسيل] III أنه رابطة حاسمة بين سينثيز الأحماض الدهنية-I ومسارات سينثيز الأحماض الدهنية-II التي تنتج الأحماض الفطرية. يشارك FAS-I في تصنيع الأحماض الدهنية C 16 و C 26 . يعمل منتج C 16 acyl-CoA كركيزة لتخليق حمض الميروميكوليك بواسطة FAS-II ، بينما يشكل الحمض الدهني C 26 فرع ألفا من حمض الميكوليك النهائي. تم اقتراح MtFabH ليكون الرابط بين FAS-I و FAS-II عن طريق تحويل C14-CoA الناتج عن FAS-I إلى C 16 -AcpM ، والذي يتم توجيهه إلى دورة FAS-II.[6] وفقًا لتحليلات توازن تدفق السيليكو ، [7] mtFabH ضروري ولكن ليس وفقًا لتحليل تهجين موقع الترانسبوزون.[8] على عكس الإنزيمات الموجودة في FAS-I ، فإن إنزيمات FAS-II ، بما في ذلك mtFabH ، ولا توجد في الثدييات ، مما يشير إلى أن مثبطات هذه الإنزيمات هي خيارات مناسبة لتطوير الأدوية.
البنية والركائز

تم النشر العلمي عن البنية البلورية لـ FabH من المتفطرة السلية ، [2][9][10] المكورات العنقودية الذهبية ، [11] الإشريكية القولونية ، [12] و ثيرموس ثيرموفيليس .[13]
تم قياس النشاط التحفيزي وخصوصية الركيزة لـ mtFabH [14] ثم تم فحصها باستخدام طرق التعيين البلوري و التطفير الموتاغينيسيس .[15] وتم تحديد هياكل ecFabH المرتبطة بالركائز ، (كو-أ ، و مالونيل-كو-أ ،و كو-أ المتحللة).[16] كما تم الإبلاغ مؤخرًا عن مثبطات جديدة تم تطويرها .[17][18][19] كذلك تم في عام 2005 الإبلاغ عن هيكل متحول لــ mtFabH المعطل بواسطة الإنزيم لورويل-كو-أ.[20]
mtFabH الأصلي هو جهاز homodimer مع M r = 77 ± 25 كيلو دالتون. على الرغم من وجود تماثل بنيوي كبير بين جميع إنزيمات FabH البكتيرية التي تم تحديدها حتى الآن ، مع قناتين لربط ركائز acyl-CoA و malonyl-ACP وثلاثي محفز محفوظ (C122 ، H258 ، N289 في mtFabH) ، يحتوي mtFabH على بقايا على طول الأسيل -قناة ربط -CoA التي تختار بشكل تفضيلي الركائز ذات السلسلة الأطول التي تبلغ ذروتها باستخدام lauroyl-CoA (C 12 ). يمكن أن تتضمن استراتيجيات التثبيط القائمة على التصميم العقلاني إزاحة تنافسية للركائز أو تعطيل الموقع الحفاز. فسفرة Thr 45 ، التي تقع عند مدخل قناة الركيزة ، تمنع النشاط ، ربما عن طريق تغيير إمكانية الوصول إلى الركائز.[21]
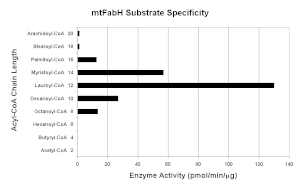
مثبطات
اثنان على الأقل من الأدوية الموجودة لمرض السل مشتق في الأصل من الميكروبات ؛ سيرولين من فطر Cephalosporium caerulens و thiolactomycin (TLM) من الفطريات الشعاعية Nocardia spp. من المعروف أن أيزونيازيد (هيدرازيد حمض أيزونيكوتينيك) ، إيثيوناميد ، تريكلوسان [ 5-كلورو -2 (2،4-ديكلوروفينوكسي) -فينول ] و TLM يثبطان على وجه التحديد تخليق حمض الميكوليك الحيوي.[22] يتم فحص مشتقات TLM والمركبات ذات الصلة لتحسين الفعالية.[23][24][25][26]
في حين تم تعلم الكثير من هذه الدراسات الهيكلية والتصميم العقلاني هو نهج ممتاز لتطوير مثبطات جديدة ، فإن الأساليب البديلة مثل التنقيب البيولوجي قد تكشف عن مركبات غير متوقعة مثل مثبط التباين الذي اكتشفه Daines et al. قد يكون هذا مهمًا بشكل خاص نظرًا لأنه يُقترح أن تكون فسفرة إنزيمات تخليق mycolate أمرًا بالغ الأهمية للتنظيم ومن المعروف أن نطاقات كيناز لديها آليات تحكم متعددة بعيدًا عن مواقع ربط ligand والمواقع النشطة.[27]
بعد اكتشاف أن أحماض الفومالينك معزولة من فطر نفايات الأوراق المعروف باسم Phoma sp. من مثبطات FabH / FabF.[28][29] وانج وآخرون. أبلغوا مؤخرًا عن اكتشافهم من بكتيريا التربة Streptomyces platensis لمثبط طبيعي جديد لـ FabH مع نشاط في الجسم الحي يسمى بلاتينسين.[30] تم العثور على هذه من خلال فحص 250000 مستخلص من بكتيريا التربة والفطريات ، مما يدل على جدوى التنقيب البيولوجي. على الرغم من كونه مضادًا حيويًا مفيدًا في حد ذاته ، فقد ثبت الآن أن بلاتنسيميسين ليس نشطًا بشكل خاص على mtFabH.[31]
من المتوقع أن تكون المثبطات الجديدة على الأرجح عبارة عن جزيئات صغيرة ذات قطبية منخفضة نسبيًا ، مع الأخذ في الاعتبار أن المواقع التحفيزية لمجانس mtFabH مخفية في جيوب كارهة للماء نسبيًا والحاجة إلى اجتياز كبسولات من العصيات المنشأة. ويدعم ذلك ضعف قابلية الذوبان في الماء لمثبط لـ ecFabH. ومن المأمول أيضًا ، من خلال كونها جزيئات صغيرة ، أن يكون تركيبها أو تخليقها الحيوي بسيطًا ورخيصًا ، مما يعزز القدرة على تحمل تكلفة الأدوية اللاحقة للبلدان النامية. تتوفر تقنيات فحص فعالية المثبطات.[32][33]
القدرة العلاجية
في عام 2005 ، تسبب مرض السل في وفاة ما يقرب من 1.6 مليون شخص في جميع أنحاء العالم ، ومرض 8.8 مليون شخص ، 90٪ من هذه الحالات في البلدان النامية ، ويقدر أن ثلث سكان العالم مصابون بالسل الكامن.[34][35] على الرغم من توفر لقاح BCG والمضادات الحيوية المتعددة ، حتى عام 2005 عاد السل إلى الظهور مرة أخرى بسبب مقاومة الأدوية المتعددة ، والتي تفاقمت بسبب حضانة ضحايا الإيدز الذين يعانون من ضعف المناعة ، وعدم الامتثال للعلاج الدوائي ، والنقص المستمر في الرعاية الصحية في البلدان النامية. يبدو أن معدلات الوفيات والإصابة بلغت ذروتها ، لكن السل لا يزال يمثل مشكلة عالمية خطيرة. هناك حاجة لعقاقير فعالة جديدة لمكافحة هذا المرض. قد يكون للمثبطات ضد mtFabH ، أو ضد الإنزيمات الأخرى لمسار FAS-II (إنزيم الحمض الدهني II)، فائدة أوسع ، مثل علاج المكورات العنقودية الذهبية المقاومة للأدوية المتعددة ، والمتصورة المنجلية ، العامل المسبب لمشكلة حرارية خطيرة أخرى ، الملاريا .
مراجع
- Davies C، Heath RJ، White SW، Rock CO (2000). "The 1.8 A crystal structure and active-site architecture of beta-ketoacyl-acyl carrier protein synthase III (FabH) from escherichia coli". Structure. ج. 8 ع. 2: 185–95. DOI:10.1016/S0969-2126(00)00094-0. PMID:10673437.
- Davies C, Heath RJ, White SW, Rock CO (2000). "The 1.8 A crystal structure and active-site architecture of beta-ketoacyl-acyl carrier protein synthase III (FabH) from escherichia coli". Structure. 8 (2): 185–95. doi:10.1016/S0969-2126(00)00094-0. PMID 10673437. نسخة محفوظة 2023-02-14 على موقع واي باك مشين.
- "Characterization of β-Ketoacyl-Acyl Carrier Protein Synthase III from Streptomyces glaucescens and Its Role in Initiation of Fatty Acid Biosynthesis". J. Bacteriol. ج. 180 ع. 17: 4481–6. 1998. DOI:10.1128/JB.180.17.4481-4486.1998. PMC:107458. PMID:9721286.
- "Identification, substrate specificity, and inhibition of the Streptococcus pneumoniae beta-ketoacyl-acyl carrier protein synthase III (FabH)". J. Biol. Chem. ج. 276 ع. 32: 30024–30. 2001. DOI:10.1074/jbc.M101769200. PMID:11375394.
- "Alteration of the Fatty Acid Profile of Streptomyces coelicolor by Replacement of the Initiation Enzyme 3-Ketoacyl Acyl Carrier Protein Synthase III (FabH)". J. Bacteriol. ج. 187 ع. 11: 3795–9. 2005. DOI:10.1128/JB.187.11.3795-3799.2005. PMC:1112031. PMID:15901703.
- "The Mycobacterium tuberculosis FAS-II condensing enzymes: their role in mycolic acid biosynthesis, acid-fastness, pathogenesis and in future drug development". Mol. Microbiol. ج. 64 ع. 6: 1442–54. يونيو 2007. DOI:10.1111/j.1365-2958.2007.05761.x. PMID:17555433.
- "Flux Balance Analysis of Mycolic Acid Pathway: Targets for Anti-Tubercular Drugs". PLOS Comput. Biol. ج. 1 ع. 5: e46. أكتوبر 2005. Bibcode:2005PLSCB...1...46R. DOI:10.1371/journal.pcbi.0010046. PMC:1246807. PMID:16261191.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - "Genes required for mycobacterial growth defined by high density mutagenesis". Mol. Microbiol. ج. 48 ع. 1: 77–84. أبريل 2003. DOI:10.1046/j.1365-2958.2003.03425.x. PMID:12657046.
- ببب: 1HND, 1HNH, 1HNJ; "Refined structures of beta-ketoacyl-acyl carrier protein synthase III". J. Mol. Biol. ج. 307 ع. 1: 341–56. مارس 2001. DOI:10.1006/jmbi.2000.4457. PMID:11243824.
- ببب: 1HZP; "Crystal structure of the Mycobacterium tuberculosis beta-ketoacyl-acyl carrier protein synthase III". J. Biol. Chem. ج. 276 ع. 23: 20516–22. يونيو 2001. DOI:10.1074/jbc.M010762200. PMID:11278743.
- ببب: 1ZOW; "Crystal structure and substrate specificity of the β-ketoacyl-acyl carrier protein synthase III (FabH) from Staphylococcus aureus". Protein Sci. ج. 14 ع. 8: 2087–94. أغسطس 2005. DOI:10.1110/ps.051501605. PMC:2279320. PMID:15987898.
- ببب: 1HN9; "Crystal structure of beta-ketoacyl-acyl carrier protein synthase III. A key condensing enzyme in bacterial fatty acid biosynthesis". J. Biol. Chem. ج. 274 ع. 51: 36465–71. ديسمبر 1999. DOI:10.1074/jbc.274.51.36465. PMID:10593943.
- ببب: 1UB7Inagaki E, Kuramitsu S, Yokoyama S, Miyano M, Tahirov TH (2007) The Crystal Structure of Beta-Ketoacyl-[Acyl Carrier Protein] Synthase III (Fabh) from Thermus thermophilus.
- "Identification and substrate specificity of beta -ketoacyl (acyl carrier protein) synthase III (mtFabH) from Mycobacterium tuberculosis". J. Biol. Chem. ج. 275 ع. 36: 28201–7. سبتمبر 2000. DOI:10.1074/jbc.M003241200. PMID:10840036.
- ببب: 1M1M, 2AJ9; "Probing the mechanism of the Mycobacterium tuberculosis beta-ketoacyl-acyl carrier protein synthase III mtFabH: factors influencing catalysis and substrate specificity". J. Biol. Chem. ج. 280 ع. 37: 32539–47. سبتمبر 2005. DOI:10.1074/jbc.M413216200. PMID:16040614.
- ببب: 1HND, 1HNH, 1HNJ; "Refined structures of beta-ketoacyl-acyl carrier protein synthase III". J. Mol. Biol. ج. 307 ع. 1: 341–56. مارس 2001. DOI:10.1006/jmbi.2000.4457. PMID:11243824.Qiu X, Janson CA, Smith WW, Head M, Lonsdale J, Konstantinidis AK (March 2001).
- ببب: 1MZS; "First X-ray cocrystal structure of a bacterial FabH condensing enzyme and a small molecule inhibitor achieved using rational design and homology modeling". J. Med. Chem. ج. 46 ع. 1: 5–8. يناير 2003. DOI:10.1021/jm025571b. PMID:12502353.
- "Structure-based design, synthesis, and study of potent inhibitors of beta-ketoacyl-acyl carrier protein synthase III as potential antimicrobial agents". J. Med. Chem. ج. 48 ع. 5: 1596–609. مارس 2005. DOI:10.1021/jm049141s. PMID:15743201.
- "A combined approach of docking and 3D QSAR study of beta-ketoacyl-acyl carrier protein synthase III (FabH) inhibitors". Bioorg. Med. Chem. ج. 14 ع. 5: 1474–82. مارس 2006. DOI:10.1016/j.bmc.2005.10.001. PMID:16275103.
- ببب: 1U6S; "Crystal structure of a substrate complex of Mycobacterium tuberculosis beta-ketoacyl-acyl carrier protein synthase III (FabH) with lauroyl-coenzyme A". J. Mol. Biol. ج. 346 ع. 5: 1313–21. مارس 2005. DOI:10.1016/j.jmb.2004.12.044. PMID:15713483.
- "The Mycobacterium tuberculosis β-Ketoacyl-Acyl Carrier Protein Synthase III Activity Is Inhibited by Phosphorylation on a Single Threonine Residue". J. Biol. Chem. ج. 284 ع. 10: 6414–24. مارس 2009. DOI:10.1074/jbc.M806537200. PMC:2649087. PMID:19074144.
- "Drugs that inhibit mycolic acid biosynthesis in Mycobacterium tuberculosis". Curr Pharm Biotechnol. ج. 3 ع. 3: 197–225. سبتمبر 2002. DOI:10.2174/1389201023378328. PMID:12164478.
- "Biphenyl-based analogues of thiolactomycin, active against Mycobacterium tuberculosis mtFabH fatty acid condensing enzyme". Bioorg. Med. Chem. Lett. ج. 13 ع. 21: 3685–8. نوفمبر 2003. DOI:10.1016/j.bmcl.2003.08.015. PMID:14552758.
- "Acetylene-based analogues of thiolactomycin, active against Mycobacterium tuberculosis mtFabH fatty acid condensing enzyme". Bioorg. Med. Chem. Lett. ج. 14 ع. 2: 373–6. يناير 2004. DOI:10.1016/j.bmcl.2003.10.061. PMID:14698162.
- "1,2-Dithiole-3-Ones as Potent Inhibitors of the Bacterial 3-Ketoacyl Acyl Carrier Protein Synthase III (FabH)". Antimicrob. Agents Chemother. ج. 48 ع. 8: 3093–102. أغسطس 2004. DOI:10.1128/AAC.48.8.3093-3102.2004. PMC:478545. PMID:15273125.
- Todd، Matthew H.، المحرر (2009). "Identification of 2-Aminothiazole-4-Carboxylate Derivatives Active against Mycobacterium tuberculosis H37Rv and the β-Ketoacyl-ACP Synthase mtFabH". PLOS ONE. ج. 4 ع. 5: e5617. Bibcode:2009PLoSO...4.5617A. DOI:10.1371/journal.pone.0005617. PMC:2680598. PMID:19440303.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - "The Mycobacterium tuberculosis β-Ketoacyl-Acyl Carrier Protein Synthase III Activity Is Inhibited by Phosphorylation on a Single Threonine Residue". J. Biol. Chem. ج. 284 ع. 10: 6414–24. مارس 2009. DOI:10.1074/jbc.M806537200. PMC:2649087. PMID:19074144.Veyron-Churlet R, Molle V, Taylor RC, Brown AK, Besra GS, Zanella-Cléon I, Fütterer K, Kremer L (March 2009).
- "Discovery of FabH/FabF Inhibitors from Natural Products". Antimicrob. Agents Chemother. ج. 50 ع. 2: 519–26. فبراير 2006. DOI:10.1128/AAC.50.2.519-526.2006. PMC:1366929. PMID:16436705.
- "Discovery of bacterial fatty acid synthase inhibitors from a Phoma species as antimicrobial agents using a new antisense-based strategy". J. Nat. Prod. ج. 69 ع. 3: 377–80. مارس 2006. DOI:10.1021/np050416w. PMID:16562839.
- "Discovery of platencin, a dual FabF and FabH inhibitor with in vivo antibiotic properties". Proceedings of the National Academy of Sciences of the United States of America. ج. 104 ع. 18: 7612–6. مايو 2007. Bibcode:2007PNAS..104.7612W. DOI:10.1073/pnas.0700746104. PMC:1863502. PMID:17456595.
- Ahmed، Niyaz، المحرر (2009). "Platensimycin Activity against Mycobacterial β-Ketoacyl-ACP Synthases". PLOS ONE. ج. 4 ع. 7: e6306. Bibcode:2009PLoSO...4.6306B. DOI:10.1371/journal.pone.0006306. PMC:2707616. PMID:19609444.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - "Mycolic Acid Index Susceptibility Method for Mycobacterium tuberculosis". J. Clin. Microbiol. ج. 39 ع. 7: 2642–5. يوليو 2001. DOI:10.1128/JCM.39.7.2642-2645.2001. PMC:88200. PMID:11427584.
- "Development of a scintillation proximity assay for beta-ketoacyl-acyl carrier protein synthase III". Anal. Biochem. ج. 282 ع. 1: 107–14. يونيو 2000. DOI:10.1006/abio.2000.4594. PMID:10860506.
- "The growing burden of tuberculosis: global trends and interactions with the HIV epidemic". Arch. Intern. Med. ج. 163 ع. 9: 1009–21. مايو 2003. DOI:10.1001/archinte.163.9.1009. PMID:12742798.
- "Global Tuberculosis Control 2007". World Health Organization. 2007. مؤرشف من الأصل في 2010-02-02. اطلع عليه بتاريخ 2010-01-02.
 بوابة علم الأحياء
بوابة علم الأحياء