حمض نووي ريبوزي رسول
الرنا المرسال أو الحمض النووي الريبوزي المرسال (بالإنجليزية: Messenger RNA) يرمز له بالرمز mRNA، هو عبارة عن جزيء رنا (حمض نووي ريبوزي) يحمل تسلسل نيوكليوتيدي يشكل الشيفرة الجينية التي على أساسها تتم صناعة جزيء ببتيد أو بروتين. يتم تكوين هذا النوع من الأحماض النووية بواسطة عملية النسخ الوراثي من الحمض النووي الريبوزي المنقوص الأوكسجين (الدنا، حمض نووي ريبوزي منقوص الأكسجين ) عن طريق إنزيم بوليميراز الحمض النووي الريبوزي (بوليميراز الحمض النووي الريبوزي). وتتم عملية تصنيع البروتين عند إرسال هذا الجزيء (الرنا المرسال) من النوية فيخرج إلى هيولي الخلية (السيتوبلازم) ؛ وتحديدا إلى جسيمات ال ريبوسوم، ليبدأ هذا الأخير عملية الترجمة وبناء البروتين في الخلية. و من هنا أخذ هذا النوع من الأحماض النووية الريبوزية اسم المرسال.[1]

الرنا الرسول وحيد وعديد السيسترون
يُقال بأن جزيء رنا رسول أنه وحيد السيسترون حين يحتوي على معلومة جينية تُترجم إلى سلسلة بروتين واحدة (عديد ببتيد)، وهذه هي الحالة لدى معظم جزيئات الرنا الرسول الخاصة بحقيقيات النوى.[2][3] من جهة أخرى، الرنا الرسول عديد السيسترون يحمل عدة إطارات قراءة مفتوحة (ORF)، يُترجم كل واحد منها إلى عديد ببتيد. عديدات الببتيد هذه عادة ما تكون لها وظيفة مشتركة أو وظائف متصلة (في العادة تكون نطاقات أو وحدات فرعية تشكل مركبا بروتينيا نهائيا) وتسلسلاتها المشفرة جُمِعت ويتم تنظيمها بمنطقة منظمة مشتركة، تحتوي على محفز وعامل. معظم الرنا الرسول المتواجد لدى البكتيريا والعتائق هو عديد السيسترون (انظر مشغل لاك) [2]، كما هو الحال في جينوم المتقدرة البشري.[4] ثنائي السيسترون هو رنا رسول يشفر بروتينين اثنين فقط.
الاصطناع والمعالجة والوظيفة
تبدأ رحلة الحياة القصيرة لجزيء الرنا الرسول بالانتساخ وتنتهي بالتحلل. خلال هذه الفترة، قد يخضع هذا الجزيء للمعالجة والتعديل والنقل قبل عملية الترجمة. تحتاج جزيئات الرنا المرسال الخاصة بالخلايا حقيقية النوى العديد من عمليات المعالجة والنقل على عكس بدائيات النوى. يُسمى مُعقد الرنا الرسول مع البروتينات المحيطة به بالبروتين النووي الريبوزي الرسول (آر إن بّي).[5]
النسخ
يبدأ النسخ عندما تؤخذ جزيئات الرنا من الدنا، وخلال هذه العملية، يشكل إنزيم بوليميراز الحمض النووي الريبوزي نسخةً من الجين الموجود على الدنا ليحملها على الرنا المرسال وفق الحاجة. تختلف هذه العملية قليلًا بين حقيقيات النوى وبدائيات النوى، وأحد أهم الاختلافات أن بوليميراز الرنا في بدائيات النوى يرتبط مع إنزيمات معالجة الدنا خلال عملية النسخ، لذا تحدث عمليتا النسخ والمعالجة في ذات الوقت. يمكن هذا شريط الرنا المرسال الجديد من التضاعف عبر إنتاج شريط متمم يُعرف بشريط الرنا الناقل. تعجز أشرطة الرنا الناقل عن تشكيل بنًى محددة بالاقتران القاعدي عندما تكون مجتمعةً.
يمثل هيكل الرنا الرسول الشريط المتمم للرنا الناقل، وهو يطابق في تسلسله تسلسل الكودونات المقابلة (الرامز المقابل) الذي يرتبط به شريط الدنا. يُسمى الجزيء الناتج المؤقت غير المعالج أو جزئي المعالجة بطليعة الحمض النووي الريبوزي الرسول، أو طليعة الرنا المرسال، وحالما تنتهي عملية المعالجة، يمكن تسمية الناتج بالرنا المرسال الناضج.
معالجة طليعة الرنا المرسال في حقيقيات النوى
تختلف معالجة الرنا المرسال كثيرًا بين حقيقيات النوى والبكتيريا والعتائق. يُعد الرنا المرسال في غير حقيقيات النوى ناضجًا في مضمونه بعد النسخ مباشرةً ولا يحتاج إلى معالجة إلا نادرًا، بينما تحتاج طليعة الرنا المرسال في حقيقيات النوى إلى العديد من خطوات المعالجة قبل نقلها إلى البلازما الخلوية وترجمتها بمساعدة الريبوسومات.[6]
التوصيل
تسمى عملية المعالجة الشاملة التي تخضع لها طليعة الرنا المرسال في حقيقيات النوى حتى تشكيل الرنا المرسال الناضج بتوصيل الرنا، وهي آلية تُزال فيها الإنترونات أو الأوترونات (مناطق غير مشفرة) من شريط الرنا وترتبط خلالها الإكسونات (مناطق مشفرة) معًا.
إضافة القبعة 5' (كاب 5')
تُعد القبعة 5' (تُسمى أيضًا قبعة الرنا، أو قبعة 7-ميثيل غوانوزين الرنا، أو قبعة إم7 جي رنا) نكليوتيد غوانين معدل يُضاف إلى «المقدمة» أو النهاية 5' للرنا المرسال في حقيقيات النوى مباشرةً بعد بدء الانتساخ. تتألف هذه القبعة من بقايا 7 ميثيل الغوانوزين الانتهائي الذي يرتبط بأول نكليوتيد منسوخ عبر الرابطة 5'-5'-ثلاثي الفوسفات. إن وجود هذا الجزيء أساسي لتعرف الريبوسوم على الرنا الرسول وحمايته من الإنزيم الحالة للرنا (ريبونكلياز).
تقترن إضافة القبعة بعملية النسخ، ويحدث انتساخ وظيفي مقترن بين حدثي نسخ شريط الرنا ومعالجته، وبالتالي يؤثر كل منهما على الآخر. بعد فترة قصيرة من النسخ، تُحاط النهاية 5' للرنا الرسول بمعقد صانع القبعة المرافق لبوليميراز الرنا. يحفز المعقد الإنزيمي التفاعلات الكيميائية المطلوبة لتقبيع الرنا المرسال، ويجري هذا الاصطناع ضمن تفاعل حيوي كيميائي متعدد الخطوات.[7]
التعديل
يخضع الرنا الرسول للتعديل في بعض الحالات، وتتغير مكوناته النكليوتيدية. مثال ذلك في البشر تعديل الرنا الرسول إلى رنا صميم البروتين الشحمي بي الرسول، الذي يخضع للتعديل في بعض النسج فقط. ينتج عن التعديل ظهور رامز إيقاف باكر يسبب إنتاج بروتين أقصر عند الترجمة.
التذييل بعديد الأدينيلات
يشمل التذييل بعديد الأدينيلات عملية الربط التساهمي بين جزء عديد الأدينيلات وجزيء الرنا الرسول. في العضويات حقيقية النوى، تُذيل أغلب النهايات 3' لجزيئات الرنا الرسول بعديد الأدينيل، لكن الدراسات الحديثة أظهرت أيضًا شيوع إضافة الامتدادات القصيرة من اليوريدين (إضافة قليل اليوريدين).[8]
تساعد إضافة ذيول عديد الأدينوزين أحادية الفوسفاتاز إلى الرنا الرسول وما يحيط بها من البروتينات على حمايته من التحلل بوساطة النوكليازات الخارجية.
يؤدي التذييل بعديد الأدينيلات أيضًا دورًا مهمًا في إنهاء عملية النسخ واستخراج الرنا الرسول من النواة وترجمته. يمكن أن يُذيل الرنا الرسول أيضًا بامتدادات الأدينوزين في العضويات بدائية النوى، لكن هنا تعمل الذيول عديدة الأدينوزين أحادية الفوسفاتاز على تسهيل التحلل بالنوكليازات الخارجية بدلًا من إعاقة هذه العملية.[9][10]
يحدث التذييل بعديد الأدينيلات خلال انتساخ الدنا إلى رنا و/أو بعده مباشرةً. بعد انتهاء عملية النسخ، تنشطر سلسلة الرنا المرسال عبر تفعيل معقد النوكلياز الداخلي المرافق لبوليميراز الرنا، ويُضاف 250 جزيء أدينوزين إلى النهاية الحرة 3' في موقع الشطر، ويُحفز هذا التفاعل ببولميراز عديد الأدينيلات. يمكن أن توجد عدة بدائل من عديد الأدينيلات للرنا المرسال الواحد كما هو الحال في عملية الوصل البديل.
قد تحدث أيضًا طفرات في موقع التذييل بعديد الأدينيلات، إذ تنشطر نسخة الرنا الأولية في موقع إضافة عديد الأدينوزين أحادي الفوسفاتاز، ويُضاف 100-200 جزيء أدينوزين إلى النهاية 3' في شريط الرنا. يمكن أن تتشكل بنية شاذة الطول أو ضعيفة الاستقرار من الرنا المرسال إذا طرأ تغير على هذا الموقع.
مراجع
- الجينوم البشري وأخلاقياته: جينات النوع البشري وجينات الفرد البشري؛ هاني رزق؛ 2007؛ دار الفكر، دمشق، سوريا.
- Kozak M (مارس 1983). "Comparison of initiation of protein synthesis in procaryotes, eucaryotes, and organelles". Microbiological Reviews. ج. 47 ع. 1: 1–45. PMC:281560. PMID:6343825.
- Niehrs C، Pollet N (ديسمبر 1999). "Synexpression groups in eukaryotes". Nature. ج. 402 ع. 6761: 483–7. DOI:10.1038/990025. PMID:10591207.
- Mercer TR، Neph S، Dinger ME، Crawford J، Smith MA، Shearwood AM، Haugen E، Bracken CP، Rackham O، Stamatoyannopoulos JA، Filipovska A، Mattick JS (أغسطس 2011). "The human mitochondrial transcriptome". Cell. ج. 146 ع. 4: 645–58. DOI:10.1016/j.cell.2011.06.051. PMC:3160626. PMID:21854988.
- Watson JD (22 فبراير 2013). Molecular Biology of the Gene, 7th edition. Pearson Higher Ed USA. ISBN:9780321851499.
- Choi YS، Patena W، Leavitt AD، McManus MT (مارس 2012). "Widespread RNA 3'-end oligouridylation in mammals". RNA (New York, N.Y.). ج. 18 ع. 3: 394–401. DOI:10.1261/rna.029306.111. PMC:3285928. PMID:22291204.
- Quaresma AJ، Sievert R، Nickerson JA (أبريل 2013). "Regulation of mRNA export by the PI3 kinase/AKT signal transduction pathway". Molecular Biology of the Cell. ج. 24 ع. 8: 1208–1221. DOI:10.1091/mbc.E12-06-0450. PMC:3623641. PMID:23427269.
- Cenik C، Chua HN، Zhang H، Tarnawsky SP، Akef A، Derti A، Tasan M، Moore MJ، Palazzo AF، Roth FP (أبريل 2011). "Genome analysis reveals interplay between 5'UTR introns and nuclear mRNA export for secretory and mitochondrial genes". PLOS Genetics. ج. 7 ع. 4: e1001366. DOI:10.1371/journal.pgen.1001366. PMC:3077370. PMID:21533221.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - Strässer K، Masuda S، Mason P، Pfannstiel J، Oppizzi M، Rodriguez-Navarro S، Rondón AG، Aguilera A، Struhl K، Reed R، Hurt E (مايو 2002). "TREX is a conserved complex coupling transcription with messenger RNA export". Nature. ج. 417 ع. 6886: 304–308. Bibcode:2002Natur.417..304S. DOI:10.1038/nature746. PMID:11979277. S2CID:1112194.
- Katahira J، Yoneda Y (27 أكتوبر 2014). "Roles of the TREX complex in nuclear export of mRNA". RNA Biology. ج. 6 ع. 2: 149–152. DOI:10.4161/rna.6.2.8046. PMID:19229134.
 بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علم الوراثة
بوابة علم الوراثة
| مقدمة لعلم الوراثة | 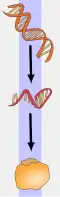 | ||||
|---|---|---|---|---|---|
| النسـخ |
| ||||
| الترجمـة |
| ||||
| مراقبة علم التخلق |
| ||||