حبيبة إجهاد
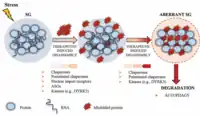
حبيبة إجهاد (بالإنجليزية: Stress granule) هي تكدسات كثيفة للبروتينات وجزيئات الرنا في العصارة الخلوية والتي تظهر حين تكون الخلية تحت الإجهاد.[1] جزيئات الرنا المخزنة هي مركبات قبل-ابتداء لترجمات معطلة: أي محاولات فاشلة لتخليق البروتين من الرنا الرسول. حجم حبيبات الإجهاد حوالي 100-200 نانومتر (عند تنقيتها بيوكيميائيا)، وهي غير محاطة بغشاء ولها صلة بالشبكة الإندوبلازمية.[2] توجد كذلك جبيبات إجهاد نووية وهذا المقال حول حبيبات الإجهاد المتكونة في العصارة الخلوية.
الوظيفة المقترحة
مازالت وظيفة جبيبات الإجهاد غير معروفة بشكل كبير، واقتُرح منذ مدة أن لها وظيفة في حماية جزيئات الرنا من الظروف المؤذية، وهذا يفسر سبب ظهورها عند الإجهاد.[3] تراكم جزيئات الرنا إلى كريات كثيفة يمكن أن يحول دون تفاعلها مع مركبات كيميائية مؤذية ويحمي المعلومة المشفَّرة في تسلسلاتها.
يمكن أن تعمل حبيبات الإجهاد كذلك كنقطة اتخاذ قرار بالنسبة لجزيئات الرنا الرسول غير المترجمة، حيث يمكن أن تمضي في أحد ثلاث مسارات: المزيد من التخزين، التفكيك أو إعادة بدء الترجمة.[4] في المقابل، تم المحاججة كذلك بأن حبيبات الإجهاد ليست مواقعا مهمة لتخزين الرنا الرسول وأنها لا تعمل كموقع وسيط لجزيئات الرنا الرسول التي هي في انتقال من حالة التخزين إلى حالة التفكيك.[5]
أظهرت مجهوداتٌ لتحديد جميع جزيئات الرنا داخل جبيبات إجهاد (نسخوم حبيبة الإجهاد) بطريقة غير منحازة عبر سَلسَلة رنا مستخلص بيوكيميائيا منها أن جزيئات الرنا لا تخزن في حبيبات الإجهاد حسب تسلسل معين، وإنما بشكل عام، مع كون الجزيئات الطويلة و/أو تلك المترجمة بشكل غير مثالي الأكثر تخزينا.[6] تشير هذه البيانات إلى أن نسخوم حبيبة الإجهاد يتأثر بتكافؤ الرنا (مع البروتينات أو جزيئات الرنا الأخرى) وبمعدل إتمام ترجمة الرنا (run-off) من قبل عديد الريبوسوم. الفرضية الأخيرة مدعومة أكثر بدراسات حديثة في تصوير الجزيء المفرد.[7] فضلا عن ذلك، قُدِّر أن حوالي 15% فقط من إجمالي الرنا الرسول في الخلية يتواجد في حبيبات الإجهاد،[6] وهذا يشير إلى أن حبيبات الإجهاد لا تؤثر سوى على مجموعة صغيرة من جزيئات الرنا الرسول في الخلية وأنها قد لا تكون ذات أهمية كبيرة في معالجة الرنا الرسول كما اعتُقد سابقا.[6][8] مع ذلك لا تمثل هذه الدراسات سوى لمحة من الزمن، ومن المحتمل أن نسبة أكبر من جزيئات الرنا الرسول تم تخزينها في مرحلة ما في حبيبات الإجهاد بسبب انتقال جزيئات الرنا من وإليها.
بروتينات الإجهاد التي هي المكون الأساسي لجبيبات الإجهاد في الخلايا النباتية هي شابرونات تعزِل وتحمي وربما ترمِّم البروتينات التي يتفكك تطويها أثناء الحر وأنواع الإجهاد الأخرى.[9][10] وعليه، أي صلة لجزيئات الرنا الرسول بحبيبات الإجهاد يمكن أن يكون ببساطة تأثيرا جانبيا لصلة البروتينات المرتبطة بالرنا المتفككة التطوي جزئيا مع حبيبات الإجهاد،[11] كما هو الحال في صلة جزيئات الرنا الرسول مع الجسيمات البروتينية.[12]
المراجع
- Gutierrez-Beltran E، Moschou PN، Smertenko AP، Bozhkov PV (مارس 2015). "Tudor staphylococcal nuclease links formation of stress granules and processing bodies with mRNA catabolism in Arabidopsis". The Plant Cell. ج. 27 ع. 3: 926–43. DOI:10.1105/tpc.114.134494. PMC:4558657. PMID:25736060.
- Kayali F، Montie HL، Rafols JA، DeGracia DJ (2005). "Prolonged translation arrest in reperfused hippocampal cornu Ammonis 1 is mediated by stress granules". Neuroscience. ج. 134 ع. 4: 1223–45. DOI:10.1016/j.neuroscience.2005.05.047. PMID:16055272.
- Nover L، Scharf KD، Neumann D (مارس 1989). "Cytoplasmic heat shock granules are formed from precursor particles and are associated with a specific set of mRNAs". Molecular and Cellular Biology. ج. 9 ع. 3: 1298–308. DOI:10.1128/mcb.9.3.1298. PMC:362722. PMID:2725500.
- Paul J. Anderson, Brigham and Women's Hospital نسخة محفوظة 22 يونيو 2006 على موقع واي باك مشين. [وصلة مكسورة]
- Mollet S، Cougot N، Wilczynska A، Dautry F، Kress M، Bertrand E، Weil D (أكتوبر 2008). "Translationally repressed mRNA transiently cycles through stress granules during stress". Molecular Biology of the Cell. ج. 19 ع. 10: 4469–79. DOI:10.1091/mbc.E08-05-0499. PMC:2555929. PMID:18632980.
- Khong A، Matheny T، Jain S، Mitchell SF، Wheeler JR، Parker R (نوفمبر 2017). "The Stress Granule Transcriptome Reveals Principles of mRNA Accumulation in Stress Granules". Molecular Cell. ج. 68 ع. 4: 808–820.e5. DOI:10.1016/j.molcel.2017.10.015. PMC:5728175. PMID:29129640.
- Khong A، Parker R (أكتوبر 2018). "mRNP architecture in translating and stress conditions reveals an ordered pathway of mRNP compaction". The Journal of Cell Biology. ج. 217 ع. 12: 4124–4140. DOI:10.1083/jcb.201806183. PMC:6279387. PMID:30322972.
- Khong A، Jain S، Matheny T، Wheeler JR، Parker R (مارس 2018). "Isolation of mammalian stress granule cores for RNA-Seq analysis". Methods. ج. 137: 49–54. DOI:10.1016/j.ymeth.2017.11.012. PMC:5866748. PMID:29196162.
- Forreiter C، Kirschner M، Nover L (ديسمبر 1997). "Stable transformation of an Arabidopsis cell suspension culture with firefly luciferase providing a cellular system for analysis of chaperone activity in vivo". The Plant Cell. ج. 9 ع. 12: 2171–81. DOI:10.1105/tpc.9.12.2171. PMC:157066. PMID:9437862.
- Löw D، Brändle K، Nover L، Forreiter C (سبتمبر 2000). "Cytosolic heat-stress proteins Hsp17.7 class I and Hsp17.3 class II of tomato act as molecular chaperones in vivo". Planta. ج. 211 ع. 4: 575–82. DOI:10.1007/s004250000315. PMID:11030557.
- Stuger R، Ranostaj S، Materna T، Forreiter C (مايو 1999). "Messenger RNA-binding properties of nonpolysomal ribonucleoproteins from heat-stressed tomato cells". Plant Physiology. ج. 120 ع. 1: 23–32. DOI:10.1104/pp.120.1.23. PMC:59255. PMID:10318680.
- Schmid HP، Akhayat O، Martins De Sa C، Puvion F، Koehler K، Scherrer K (يناير 1984). "The prosome: an ubiquitous morphologically distinct RNP particle associated with repressed mRNPs and containing specific ScRNA and a characteristic set of proteins". The EMBO Journal. ج. 3 ع. 1: 29–34. DOI:10.1002/j.1460-2075.1984.tb01757.x. PMC:557293. PMID:6200323.
 بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي