جهاز غولجي
جهاز غولجي أو معقد غولجي أو جسيم غولجي أو جسم غولجي أو شبكة غولجي (غولجي تكتب أيضاً جولجي) (بالإنجليزية: Golgi apparatus)[1][2] هي عضية خلوية تتواجد في معظم خلايا حقيقيات النوى،[3] اكتشفها العالم الإيطالي كاميلو غولجي سنة 1897 وسماها باسمه سنة 1898.[4]
| علم الأحياء الخلوي | |
|---|---|
| الخلية الحيوانية | |
 مكونات الخلية الحيوانية النموذجية:
| |
| جهاز جولجي | |
|---|---|
 | |
| المكتشف | كاميلو جولجي |
| سنة الاكتشاف | 1898 |
| الطول | 0.5 - 2 ميكرون |
| التواجد | الحيوانات النباتات الفطريات الطلائعيات |
| الموقع | بين النواة والغشاء البلازمي للخلية |
جهاز غولجي جزء من النظام الغشائي الداخلي في السيتوبلازم، ويتكون من عدة حزمات صهريجية متصلة ببعضها عبر أنيبيبات دقيقة ويقوم بمعالجة وتجميع البروتينات في حويصلات غشائية ناقلة وإرسالها إلى وجهاتها النهائية. يقع جهاز غولجي لدى الثدييات في تقاطع المسارات الإفرازية واليحلولية ومسار الإدخال بجوار نواة الخلية بالقرب من الجسيم المركزي وله أهمية خاصة في معالجة البروتينات من أجل إفرازها، ويحتوي على مجموعة من إنزيمات الغلكزة التي تضيف عدة موحودات سكرية إلى البروتينات أثناء مرورها داخل الجهاز.
الاكتشاف

بسبب حجمه الكبير وبنيته المميزة، كان جهاز غولجي من أول العضيات التي تم اكتشافها وملاحظتها بدقة، اكتُشف جهاز غولجي سنة 1898 بواسطة الفيزيائي الإيطالي كاميلو غولجي أثناء بحوثه في الجهاز العصبي.[4][5] بعد مشاهدة الجهاز أول مرة بالمجهر الضوئي سماه «جهاز الشبكة الداخلية»، وشكك البعض في اكتشافه في ذلك الوقت، محاججين بأن ظهور البنية لم يكن سوى وهما بصريا أحدثته تقنية الملاحظة التي استخدمها غولجي، ومع تطور المجاهر الضوئية الحديثة في القرن الـ20 تم تأكيد الاكتشاف.[6] الإشارات الأولية إلى جهاز غولجي كانت بأسماء متعددة منها: «جهاز غولجي-هولمغرن»، «قنوات غولجي-هولمغرن» و«جهاز غولجي-كوبش».[4] استُخدم مصطلح «جهاز غولجي» سنة 1910 وظهر أول مرة في منشورات علمية سنة 1913، في حين أن مصطلح «معقد غولجي» استخدم سنة 1956.[4]
الموقع الخلوي
يختلف موقع جهاز غولجي بين حقيقيات النوى، حيث يتواجد لدى الثدييات جهاز غولجي واحد يقع بجوار نواة الخلية بالقرب من الجسيم المركزي. روابط أنبوبية هي المسؤولة عن ربط الحُزَمِ معا، تموقع جهاز غولجي وأنابيب الربط الخاصة به يعتمد على الأنيبيبات الدقيقة، وقد أظهرت تجاربٌ أنه حين تتفكك الأنيببات الدقيقة يفقد جهاز غولجي روابطه المشتركة وتصبح الحزم منفردة ومتوزعة في السيتوبلازم.[7] يتواجد لدى الخميرة العديد من أجهزة غولجي منتشرة في السيتوبلازم (كما لوحظ لدى فطريات الخميرة)، حُزم غولجي لدى النباتات ليست مركَّزَة في منطقة الجسيم المركزي ولا تشكل شرائط غولجي.[8] يعتمد تنظيم جهاز غولجي لدى النباتات على خيوط الأكتين وليس الأنيببات الدقيقة.[8] الميزة المشتركة بين أجهزة غولجي هي أنها مجاورة لمواقع الخروج في الشبكة الإندوبلازمية.[9]
البنية
_ar.svg.png.webp)
يتكون جهاز غولجي لدى معظم حقيقيات النوى من سلسلة من الأحياز وهو مجموعة من الفجوات الغشائية المسطحة المدمجة وتعرف باسم الصهاريج (وتسمى كذلك بالجسيم المشتبك)، نشأت من تكدسات حويصلات تتبرعم من الشبكة الإندوبلازمية. تحتوي خلايا الثديات عادة على حوالي 40 إلى 100 حزمة من الصهاريج.[10] حيث تتواجد من أربع إلى ثمانية صهاريج في كل حزمة، لكن لدى بعض الطلائعيات تمت ملاحظة ما يصل إلى ستين صهريجا.[6] تنقسم مجموعة الصهاريج هذه إلى ثلاث أحياز: مقرون، وسط ومفروق مشكلين شبكتين أساسيتين شبكة مقرون غولجي (CGN) وشبكة مفروق غولجي. (TGN) شبكة مقرون هي أول صهريج وشبكة مفروق هي آخر صهريج والذي تُجمّع فيه البروتينات داخل الحويصلات التي تغادر إلى اليحلولات، الحويصلات الإفرازية أو سطح الخلية. عادة ما تكون شبكة مفروق متموضعة بجوار الحزمة، لكن يمكن أن تكون منفصلة عنها. الشبكة مفروق يمكن أن تتصرف كدخلول سابق لدى الخميرة والنباتات.[8]
توجد اختلافات بنيوية وتنظيمية في جهاز غولجي بين حقيقيات النوى، فلدى بعض الخميرات تحزيم غولجي (تحزيم الصهاريج في مجموعة) غير مُلاحظ، فمثلا الخميرة بيكيا باستوريس لديها جهاز غولجي محزَّم بينما السكيراء الجعوية ليس لديها جهاز محزم. في النباتات، يظهر أن الحزمات الفردية لجهاز غولجي تعمل بشكل منفرد.[8] يميل جهاز غولجي لأن يكون أكبر حجما وعددا في الخلايا التي تُخلِّق وتطرح كميات كبيرة من المركبات، على سبيل المثال الخلية البلازمية باء التي تفرز الأجسام المضادة الخاصة بالجهاز المناعي لديها معقدات غولجي كثيرة.
في جميع حقيقيات النوى، لدى كل حزمة من الصهاريج مدخل مقرون (سيس) ومخرج مفروق (ترانس). تتميز هذه الوجوه ببنية وكيميائية حيوية فريدة.[11] توجد داخل الحزم الفردية تشكيلات من الإنزيمات المسؤولة على اختيار وتعديل البروتينات وشحنها، حيث تؤثر هذه التعديلات على مصير البروتين ووظيفته. تقسيم أحياز جهاز غولجي له ميزات في فصل الإنزيمات عن بعضها وبذلك الحفاظ خطوات معالجة البروتينات الاختيارية والمتواصلة: بحيث تُجمع الإنزيمات المحفِّزة للتعديلات الأولية في الوجه مقرون للصهاريج، أما الإنزيمات التي تحفز التعديلات اللاحقة أو الأخيرة فتوجد في الوجه مفروق لصهاريج حزمة جهاز غولجي.[7][11]
الوظيفة

يقوم جهاز غولجي بمعالجة البروتينات القادمة من الشبكة الإندوبلازمية وتجميعها وشحنها إلى وجهاتها المختلفة داخل وخارج الخلية. تُجمّع البروتينات المخلقة في الشبكة الإندوبلازمية في حويصلات وترسل إلى جهاز غولجي حيث تندمج معه، تتعرض شحنات البروتين هذه إلى التعديل والمعالجة ثم تشحن مجددا لإفرازها خارج الخلية عبر الإخراج الخلوي أو لاستخدامها في أماكن مختلفة من الخلية. وعليه يمكن اعتبار جهاز غولجي كمركز نقل وتوزيع: يقوم بوسم الشحنات وإرسالها إلى أماكن مختلفة من الخلية أو إلى المحيط خارج الخلوي. يقوم جهاز غولجي كذلك بنقل الليبيدات وتكوين اليحلولات.[12]
لبنية جهاز غولجي صلة وثيقة بوظيفته. لدى الحُزم الفردية تشكيلات مختلفة من الإنزيمات وهذا يسمح بمعالجة تقدُمِيّة لشحنات البروتين أثناء انتقالها داخل الصهاريج إلى الوجه مفروق.[7][11] تحدث التفاعلات الإنزيمية داخل حزم غولجي بشكل حصري بالقرب من سطح أغشيتها حيث توجد الإنزيمات مدمجة، وهذا عكس الشبكة الإندوبلازمية التي تملك بروتينات قابلة للذوبان وإنزيمات في لمعتها. معظم المعالجة الإنزيمية هي تعديلات ما بعد الترجمة تخضع لها البروتينات، على سبيل المثال: تحدث فسفرة قليلات السكاريد في البروتينات اليحلولية في أول الوجه مقرون CGN،[7] كما يتم في الصهريج مقرون نزع جزيئات المانوز.[7][11] وتتم إزالة جزيئات المانوز وإضافة N-أسيتيل غلوكوز أمين في الصهاريج الوسطية.[7] تحدث إضافة الجلاكتوز وحمض السياليك في الصهاريج مفروق.[7] تحدث كبرتة التيروسين والكاربوهيدرات كذلك في الوجه مفروق. تعديلات ما بعد الترجمة الأخرى للبروتينات تشمل إضافة الكاربوهيدرات (غلكزة) [13] والفوسفات (فسفرة).قد يتم في تعديلات ما بعد الترجمة إضافة تسلسلٍ يحدد الوجهة الأخيرة للبروتين، فمثلا يضيف جهاز غولجي وسم مانوز 6-فوسفات للبروتينات التي وجهتها اليحلولات. وظيفة أخرى مهمة يقوم بها جهاز غولجي هي تكوين البروتيوغليكانات، إذ توجد إنزيمات في الجهاز تقوم بإضافة بروتيناتٍ إلى الغليكوز أمينوغليكانات مشكلة بروتيوغليكانات.[14] الغليكوز أمينوغليكانات هي جزيئات عديدات سكاريد طويلة غير متفرعة متواجدة في النسيج البيني خارج الخلية لدى الحيوانات.
النقل الحويصلي

تقصد الحويصلات التي تغادر الشبكة الإندوبلازمية الوجه مقرون لجهاز غولجي، أين تندمج مع غشاء غولجي وتُفرغ حمولتها في لمعته. حين تدخل الجزيئات اللمعة يتم تعديلها ثم تنصيفها وشحنها لنقلها إلى وجهاتها التالية.
تنتقل البروتينات التي لها وجهة غير الشبكة الإندوبلازمية وجهاز غولجي عبر صهاريج غولجي نحو الوجه مفروق، إلى شبكة معقدة من الأغشية والحويصلات تعرف بوجه مفروق-غولجي (TGN). في هذه المكان من الجهاز يتم تصنيف البروتينات وشحنها إلى وجهاتها النهائية عبر وضعها في إحدى ثلاثة أنواع من الحويصلات وذلك حسب الببتيد الإشعاري الذي تحمله.
| النوع | وصف | مثال |
|---|---|---|
| حويصلات إيماسية (تكوينية) | حويصلات تحتوي على بروتينات وجهتها خارج الخلية. بعد التجميع، تتبرعم الحويصلات وتنتقل مباشرة نحو الغشاء البلازمي وتندمج معه محررة محتواها في المحيط خارج الخلوي في عملية تعرف بالإفراز. | الأجسام المضادة المفرزة من قبل الخلايا البلازمية البائية. |
| حويصلات إفرازية (منظمة) | حويصلات تحتوي على بروتينات وجهتها خارج الخلية. بعد التجميع، تتبرعم الحويصلات ويتم تخزينها في الخلية حتى تُعطى إشارة لإفرازها. عند استقبال هذه الإشارة تتحرك الحويصلات نحو الغشاء وتندمج معه لتحرير حمولتها. تعرف هذه العملية بالإفراز المنظم. | النواقل العصبية المفرزة من قبل العصبونات. |
| حويصلات يحلولية | حويصلات تحتوي على بروتينات وريبوسومات وجهتها اليحلول وهو عضية محلِّلة تحتوى على العديد من الهيدرولازات الحمضية، أو عضيات تخزين مشابهة لليحلول. تشمل هذه البروتينات كلا من البروتينات الهضمية والغشائية. تندمج الحويصلة أولا مع الدخلول ثم ينتقل المحتوى إلى اليحلول عبر آليات غير معروفة. | البروتياز الهضمي الذي وجهته اليحلول. |
بريفيلدين A
بريفيلدين A (BFA) هو مستقلب فطري يُسستخدم تجريبيا لتعطيل المسار الإفرازي وكطريقة لاختبار وظيفة جهاز غولجي.[15] يثبط البريفديلين A تنشيط بعض عوامل ربوزة الـADP (ARF).[16] عوامل الربوزة هذه عبارة عن غتبازات صغيرة تعمل على تنظيم النقل الحويصلي بواسطة ترابط بروتينات COP بالدخلولات وبجهاز خولجي.[16] كما يثبط البريفيلدين A عمل عدة عوامل تبديل نوكليوتيد الغوانين (GEFs) والتي تتوسط ارتباط عوامل الربوزة (ARFs) بالغوانوزين ثلاثي الفوسفات. معالجة الخلية بالبريفيلدين A يعطل المسار الإفرازي ويحفز تفكيك جهاز غولجي وتوزيع بروتيناته على الدخلولات والشبكة الإندوبلازمية.[15][16]
معرض الصور
- ديناميكية غولجي في الخميرة، الأخضر وسم لغولجي الأولي والأحمر وسم لغولجي الناضج.[17]
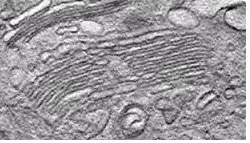 حزمتي غولجي متصلتين كشريط في خلية فأر، مأخوذة من الفيديو
حزمتي غولجي متصلتين كشريط في خلية فأر، مأخوذة من الفيديو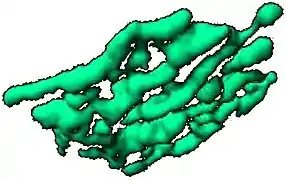 إسقاط ثلاثي الأبعاد لحزمة غولجي خاصة بالثدييات مصورة بالمجهر البؤري، ورسم حجم السطح تم ببرنامج Imaris، مأخوذة من الفيديو
إسقاط ثلاثي الأبعاد لحزمة غولجي خاصة بالثدييات مصورة بالمجهر البؤري، ورسم حجم السطح تم ببرنامج Imaris، مأخوذة من الفيديو
المصادر
- apparatus&field_magal=Medical "Al-Qamoos القاموس - English Arabic dictionary / قاموس إنجليزي عربي". مؤرشف من الأصل في 2017-12-03. اطلع عليه بتاريخ 2016-12-21.
{{استشهاد ويب}}: تحقق من قيمة|مسار أرشيف=(مساعدة) - "ترجمة و معنى كلمة golgi apparatus في قاموس المعاني. قاموس عربي انجليزي مصطلحات صفحة 1". مؤرشف من الأصل في 2018-11-21. اطلع عليه بتاريخ 2016-12-21.
- Pavelk M، Mironov AA (2008). The Golgi Apparatus: State of the art 110 years after Camillo Golgi's discovery. Berlin: Springer. ص. 580. ISBN:978-3-211-76310-0. مؤرشف من الأصل في 2019-03-21.
- Fabene PF، Bentivoglio M (1998). "1898–1998: Camillo Golgi and "the Golgi": one hundred years of terminological clones". Brain Res. Bull. ج. 47 ع. 3: 195–8. DOI:10.1016/S0361-9230(98)00079-3. PMID:9865849.
- Golgi C (1898). "Intorno alla struttura delle cellule nervose" (PDF). Bollettino della Società Medico-Chirurgica di Pavia. ج. 13 ع. 1: 316. مؤرشف من الأصل (PDF) في 2018-04-07.
- Davidson MW (13 ديسمبر 2004). "The Golgi Apparatus". Molecular Expressions. Florida State University. مؤرشف من الأصل في 2006-11-07. اطلع عليه بتاريخ 2010-09-20.
- Alberts، Bruce؛ وآخرون. Molecular Biology of the Cell. Garland Publishing. ISBN:978-0-8153-1619-0. مؤرشف من الأصل في 2009-09-11.
- Nakano A، Luini A (2010). "Passage through the Golgi". Curr Opin Cell Biol. ج. 22 ع. 4: 471–8. DOI:10.1016/j.ceb.2010.05.003. PMID:20605430.
- Suda Y، Nakano A (2012). "The yeast Golgi apparatus". Traffic. ج. 13 ع. 4: 505–10. DOI:10.1111/j.1600-0854.2011.01316.x. PMID:22132734.
- Duran JM، Kinseth M، Bossard C، Rose DW، Polishchuk R، Wu CC، Yates J، Zimmerman T، Malhotra V (2008). "The role of GRASP55 in Golgi fragmentation and entry of cells into mitosis". Mol. Biol. Cell. ج. 19 ع. 6: 2579–87. DOI:10.1091/mbc.E07-10-0998. PMC:2397314. PMID:18385516.
- Day KJ، Staehelin LA، Glick BS (2013). "A three-stage model of Golgi structure and function". Histochem. Cell Biol. ج. 140 ع. 3: 239–49. DOI:10.1007/s00418-013-1128-3. PMC:3779436. PMID:23881164.
- Campbell، Neil A (1996). Biology (ط. 4). Menlo Park, CA: Benjamin/Cummings. ص. 122, 123. ISBN:0-8053-1957-3.
- William G. Flynne (2008). Biotechnology and Bioengineering. Nova Publishers. ص. 45–. ISBN:978-1-60456-067-1. مؤرشف من الأصل في 2020-01-03. اطلع عليه بتاريخ 2010-11-13.
- Prydz K، Dalen KT (يناير 2000). "Synthesis and sorting of proteoglycans". J. Cell Sci. 113. ج. Pt 2: 193–205. PMID:10633071.
- Marie M، Sannerud R، Avsnes Dale H، Saraste J (2008). "Take the 'A' train: on fast tracks to the cell surface". Cell Mol Life Sci. ج. 65 ع. 18: 2859–74. DOI:10.1007/s00018-008-8355-0. PMID:18726174.
- D'Souza-Schorey C، Chavrier P (2006). "ARF proteins: roles in membrane traffic and beyond". Nat Rev Mol Cell Biol. ج. 7 ع. 5: 347–58. DOI:10.1038/nrm1910. PMID:16633337.
- Papanikou E، Day KJ، Austin J، Glick BS (2015). "COPI selectively drives maturation of the early Golgi". eLife. ج. 4. DOI:10.7554/eLife.13232. PMC:4758959. PMID:26709839. مؤرشف من الأصل في 2017-10-03.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link)
 بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي