ترابط هوغستين
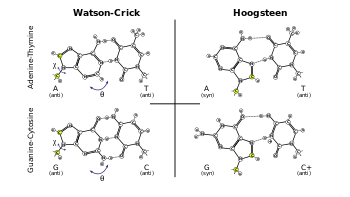
ترابط قواعد هوغستين هو نوع آخر من ترابط القواعد في الأحماض النووية، يمكن في هذا النوع ربط قاعدتين نوويتين من كلا السلسلتين بواسطة روابط هيدروجينية في الثلم الكبير. زوج قواعد هوغستين يستخدم الذرة N7 في البيورين كمستقبل للرابطة الهيدروجينية وذرة الكربون C6 المرتبطة بمجموعة الأمين كمانح، ويرتبط البيورين بالبيريميدين في الوجه (N3–C4) كما في حالة واتسون كريك العادية.
تاريخ
بعد عشر سنوات من نشر جيمس واتسون وفرنسيس كريك نموذجهما للولب الدنا المزدوج [2] أشار كارست هوغستين [3] إلى وجود بنية كريستالية لمركب قامت فيه النظائر A وT بتشكيل زوج قاعدي يختلف في هندسته الفراغية عن ذلك الذي أشار إليه واتسون وكريك، وأشار إلى وجود زوج قاعدي G≡C آخر مختلف كذلك. أبرز هوغستين أن وجود هذا الترابط الهيدروجيني البديل في الدنا يشير إلى احتمال وجود شكل مختلف للدنا، مع ذلك فإن ترابط قواعد هوغستين نادر الملاحظة.
الخصائص الكيميائية
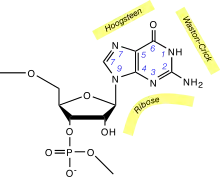
لترابط هوغستين خصائص مختلفة عن واتسون كريك، الزاوية بين الرابطتين الغلوسيديتين (حوالي 80° في الزوج A=T) أكبر والمسافة C1′–C1′ (860 بيكومتر، أو 8.6 أنغستروم) أصغر من الهندسة العادية لدى واتسون كريك، في بعض الحالات التي تكون فيها جزيئتا السكر على جانبين مختلفين من محور الزوج القاعدي يسمى ذلك ترابط هوغستين العكسي وتكون فيه إحدى القاعدتين ملتفة 180° بالنسبة للأخرى.
تتواجد أزواج قواعد هوغستين في بعض تسلسللات الدنا وخاصة النوكليوتيدات الثنائية CA وTA ككيانات مؤقتة موجودة في التوازن الحراري مع أزواج قواعد واتسون كريك قياسية، يتطلب كشف الأنواع المؤقتة استخدام تقنيات مطيافية الرنين المغناطيسي النووي التي لم يتم تطبيقها على الجزيئات الضخمة إلا حديثا.[1]، تمت ملاحظة ترابط هوغستين في مركبات الدنا والبروتين [4]، تطورت بعض البروتينات للتعرف على زوج قاعدي واحد فقط وتستخدم تفاعلات بين جزيئية لإزاحة التوازن بين كلا الهندستين.
للدنا ميزات عديدة تسمح لتسلسلات محددة بأن تتعرف عليها البروتينات، اعتُقد في البداية أن هذا التعرف يتعلق بتفاعلات ارتباط براوبط هيدروجينية محددة بين السلاسل الجانبية للحمض الأميني والقواعد، لكن اتضح سريعا أنه لا توجد علاقات وأنماط محددة يمكن التعرف عليها (لا يوجد كود بسيط يمكن قراءته) وذلك لأن الدنا يمكن أن يخضع لتغيرات بنيوية تغير من شكل اللولب المزدوج التقليدي، وتُغير البنى الناتج عرض قواعد الدنا لجزيئات البروتينات وذلك يؤثر في آلية التعرف.
لأن تغيرات بنية اللولب المزدوج نفسها تعتمد على تسلسل القواعد، يمكن للبروتينات التعرف على الدنا بطريقة مماثلة للطريقة التي تتعرف بها على البروتينات الأخرى وجزيئات الربيطات الصغيرة الأخرى، كمثال عبر الشكل الهندسي الفراغي (بدل تسلسل محدد). على سبيل المثال امتدادات قواعد A وT يقود إلى تضييق الثلم الصغير في الدنا ما ينتج عنة إمكانيات كهروستاتية سلبية معززة تنشأ مواقع ارتباط لجزيئات حمض الأمين أرجنين الموجب الشحنة المتواجدة في البروتين.
مراجع
- Evgenia N. Nikolova؛ Eunae Kim؛ Abigail A. Wise؛ Patrick J. O'Brien؛ Ioan Andricioaei؛ Hashim M. Al-Hashimi (2011). "Transient Hoogsteen base pairs in canonical duplex DNA". Nature. ج. 470: 498–502. Bibcode:2011Natur.470..498N. DOI:10.1038/nature09775. PMC:3074620.
- Watson JD، Crick FH (1953). "Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid". Nature. ج. 171 ع. 4356: 737–738. Bibcode:1953Natur.171..737W. DOI:10.1038/171737a0. PMID:13054692.
- Hoogsteen K (1963). "The crystal and molecular structure of a hydrogen-bonded complex between 1-methylthymine and 9-methyladenine". Acta Crystallographica. ج. 16: 907–916. DOI:10.1107/S0365110X63002437.
- Jun Aishima, Rossitza K. Gitti, Joyce E. Noah, Hin Hark Gan, Tamar Schlick, Cynthia Wolberger (2002). "A Hoogsteen base pair embedded in undistorted B‐DNA". Nucleic Acids Res. ج. 30 ع. 23: 5244–5252. DOI:10.1093/nar/gkf661.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: أسماء متعددة: قائمة المؤلفين (link)
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية