بروتين سيكلين دي
سيكلين دي هو عضو في عائلة بروتين يشارك في تنظيم تقدم دورة الخلية. يبدأ توليف سيكلين دي أثناء جي ون ويدفع انتقال المرحلة جي ون على اس. بروتين سيكلين دي في أي مكان من 155 (في زيبرا بلح البحر) إلى 477 (في ذبابة الفاكهة) الأحماض الأمينية في الطول.
| cyclin D1 | |
|---|---|
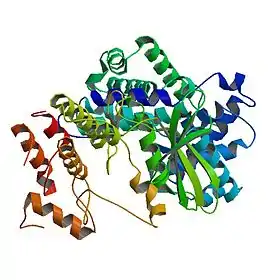 Crystal structure of human cyclin D1 (blue/green) in complex with cyclin-dependent kinase 4 (yellow/red).[1] | |
| المعرفات | |
| الرمز | CCND1 |
| رموز بديلة | BCL1, D11S287E, PRAD1 |
| أنتريه | 595 |
| HUGO | 1582 |
| أوميم | 168461 |
| RefSeq | NM_053056 |
| يونيبروت | P24385 |
| بيانات أخرى | |
| الموقع الكروموسومي | Chr. 11 q13 |
بمجرد وصول الخلايا إلى حجم الخلية الحرج (وإذا لم يكن هناك شريك تزاوج في الخميرة) وإذا كانت هناك عوامل نمو ومُحسنات (للكائنات متعددة الخلايا) أو المغذيات (للكائنات أحادية الخلية)، فإن الخلايا تدخل في دورة الخلية. بشكل عام، يتم فصل جميع مراحل دورة الخلية ترتيبًا زمنيًا في البشر ويتم تشغيلها بواسطة مجمعات سيكلين- سي دي كي التي يتم التعبير عنها بشكل دوري وتكرار جزئي في الوظيفة. سيكلين هي بروتينات حقيقية النواة التي تشكل إنزيمات مع بروتينات تعتمد على سيكلين (سي دي كي)، والتي تنشطها. يتم تنظيم وفرة السيكلانات بشكل عام عن طريق تخليق البروتين وتدهوره من خلال مسار يعتمد على مجمع تعزيز المرحلة.
سيسكلين دي هو واحد من السيكلانات الرئيسية المنتجة من حيث أهميته الوظيفية. يتفاعل مع أربعة سي دي كي اس: كيناز يعتمد على سيكلين 2 و 4 و 5 و 6 . في الخلايا المتكاثرة، يكون تراكم مركب سيكلن كيناز يعتمد على سيكلين دي 4 / 6 ذو أهمية كبيرة لتقدم دورة الخلية. على وجه التحديد، مركب سيكلن كيناز يعتمد على سيكلين دي 4 / 6 معقد جزئيًا لبروتين الورم الأرومي الأرومي الشبكي (آر بي)، والذي يمكن أن يؤدي تثبيطه إلى التعبير عن بعض الجينات (على سبيل المثال: سيكلن إي) مهم لتطور المرحلة إس.
ذبابة الفاكهة والعديد من الكائنات الحية الأخرى لديها بروتين سيكلن دي واحد فقط. في الفئران والبشر، تم تحديد بروتينين آخرين لـ سيكلين دي. يتم التعبير عن المتجانسات الثلاثة، التي تسمى سيكلين دي 1 و <a href=«./ سيكلين د 2» rel="mw:WikiLink" data-linkid="151" data-cx="{&quot;adapted&quot;:false,&quot;sourceTitle&quot;:{&quot;title&quot;:&quot;Cyclin D2&quot;,&quot;thumbnail&quot;:{&quot;source&quot;:&quot;http://upload.wikimedia.org/wikipedia/commons/thumb/e/e7/Ideogram_human_chromosome_12.svg/80px-Ideogram_human_chromosome_12.svg.png&quot;,&quot;width&quot;:80,&quot;height&quot;:32},&quot;description&quot;:&quot;protein-coding gene in the species Homo sapiens&quot;,&quot;pageprops&quot;:{&quot;wikibase_item&quot;:&quot;Q5198271&quot;},&quot;pagelanguage&quot;:&quot;en&quot;},&quot;targetFrom&quot;:&quot;mt&quot;}" class="cx-link" id="mwIg" title=«سيكلين د 2»>سيكلين دي</a>2 و <a href=«./ سيكلين د 3» rel="mw:WikiLink" data-linkid="152" data-cx="{&quot;adapted&quot;:false,&quot;sourceTitle&quot;:{&quot;title&quot;:&quot;Cyclin D3&quot;,&quot;thumbnail&quot;:{&quot;source&quot;:&quot;http://upload.wikimedia.org/wikipedia/commons/thumb/0/03/Ideogram_human_chromosome_6.svg/80px-Ideogram_human_chromosome_6.svg.png&quot;,&quot;width&quot;:80,&quot;height&quot;:32},&quot;description&quot;:&quot;protein-coding gene in the species Homo sapiens&quot;,&quot;pageprops&quot;:{&quot;wikibase_item&quot;:&quot;Q5198273&quot;},&quot;pagelanguage&quot;:&quot;en&quot;},&quot;targetFrom&quot;:&quot;mt&quot;}" class="cx-link" id="mwIw" title=«سيكلين د 3»>سيكلين دي</a> 3 في معظم الخلايا التكاثرية وتختلف المقادير النسبية المعبر عنها في أنواع الخلايا المختلفة.
المثليون
تم العثور على أكثر المتجانسات المدروسة من سيكلين دي في الخميرة والفيروسات.
يتفاعل خميرة سيكلين دي، المشار إليها باسم سي إل إن 3، مع سي دي سي 28 (بروتين التحكم في انقسام الخلايا) خلال جي1.
في الفيروسات، مثل فيروس الهربس وفيروس الهربس المرتبط بساركوما كابوزي اكتسبت سيكلين دي وظائف جديدة من أجل معالجة عملية التمثيل الغذائي للخلية المضيفة لصالح الفيروسات. يربط سيكلين دي الفيروسي سي دي كي6 البشري ويثبط آر بي عن طريق الفسفرة، مما يؤدي إلى عوامل النسخ المجانية التي تؤدي إلى نسخ البروتين الذي يعزز المرور عبر مرحلة جي1 من دورة الخلية. البعض من الروبيديوم، الفيروسية امركب سيكلن كيناز يعتمد على سيكلين دي6 المجمع أيضا يستهدف بي27 كيب، مثبط للمعلمين من السيكلين إي وأي. وبالإضافة إلى ذلك، السيكلين الفيروسي مركب سيكلن كيناز يعتمد على سيكلين دي6 مقاوم للمثبطات معلمين، مثل المثبط بي21 سي آي بي1 / واف 1 والجين بي16 إنك 4 أي التي في الإنسان تمنع الخلايا سي دي كي4 بمنعها من تكوين مركب نشط مع سيكلن د.
بناء
يمتلك سيكلين دي هيكلًا ثلاثيًا مشابهًا للأدوية الأخرى التي تسمى طية سيكلين. يحتوي هذا على نواة من مجالين مضغوطين لكل منهما خمسة حلزونات ألفا. أول حزمة خماسية الحلزون هي صندوق سيكلان محفوظ، وهي منطقة تحتوي على حوالي 100 بقايا حمض أميني في جميع السيكلينات، وهي ضرورية لربط سي دي كي وتنشيطها. تتكون حزمة اللولب الخمسة الثانية من نفس ترتيب اللوالب، لكن التسلسل الأساسي للنطاقين الفرعيين متميز. تحتوي جميع سيكلات دي من النوع دي (دي1 و دي2 و دي3) على نفس اللطخة الحلزونية ألفا 1. ومع ذلك، فإنه يتألف من بقايا مختلفة من الأحماض الأمينية مثل الرقعة نفسها في السيكلان إي وآي وبي.
وظيفة
عوامل النمو تحفز عائلة راس / راف / كينازات خارج الخلية ينظمها الاشارات التي تحفز إنتاج سيكلين دي. أحد أعضاء المسارات، ينشط بروتين كيناز منشط الميتوجين عامل النسخ بروتين ميس، الذي يغير نسخ الجينات المهمة في دورة الخلية، ومن بينها سيكلين دي. وبهذه الطريقة، يتم تصنيع سيكلين دي طالما كان عامل النمو موجودًا.
يتم الحفاظ على مستويات سيكلين دي في الخلايا المتكاثرة طالما أن عوامل النمو موجودة، فإن اللاعب الرئيسي لانتقال جي 1 على إس هو مجمعات سيكلين مركب سيكلن كيناز يعتمد على سيكلين دي4 / 6 النشطة. لا يؤثر سيكلين دي على انتقال جي1 على إس إلا إذا كان معقدًا باستخدام مركب سيكلن كيناز يعتمد على سيكلين دي 4 أو 6.
انتقال بروتين جي 1 على إس
واحدة من أفضل ركائز مركب سيكلن كيناز يعتمد على سيكلين دي4 و -6 هي بروتين الورم المثبط للورم الأرومي الشبكي آر بي هو منظم مهم للجينات المسؤولة عن التقدم خلال دورة الخلية، ولا سيما من خلال المرحلة جي 1 على إس..
يقترح أحد النماذج أن كميات سيكلين دي، وبالتالي نشاط سيكلين مركب سيكلن كيناز يعتمد على سيكلين دي4 و -6، تزداد تدريجيًا خلال جي1 بدلاً من التأرجح في نمط محدد كما تفعل إس وسيكلين إم. يحدث هذا استجابة لأجهزة استشعار الإشارات التنظيمية للنمو الخارجي ونمو الخلايا، ونتيجة لذلك يتم فسفرة آر بي. الذي يقلل من ارتباطه بـ غي 2 إف وبالتالي يسمح بالتنشيط بوساطة إي 2 إف لنسخ سيكلين إي وسيكلين أي، اللذين يرتبطان بـ مركب سيكلن كيناز يعتمد على سيكلين 1 ومركب سيكلن كيناز يعتمد على سيكلين 2 على التوالي لإنشاء مجمعات تستمر مع الفسفرة آر بي. تعمل معقدات كيناز المعتمدة من سيكلين إي وأي أيضًا على تثبيط الوحدة الفرعية البروتين الذي يقوم بتجنيد إنزيم إي إلى الركيزة البروتينية 2 إلى أي بي سي على سي التي تنشط الوحدة الفرعية سي دي اتش1 من خلال الفسفرة، والتي تعمل على تثبيت الركائز مثل سيكلين إي. تعمل الكينازات التابعة على دفع الالتزام بانقسام الخلايا إلى نقطة تفتيش جي 1 على إس وبعدها.
يقترح نموذج آخر أن مستويات سيكلين دي تظل ثابتة تقريبًا من خلال جي 1. آر بي هو أحادي الفسفرة خلال وقت مبكر إلى منتصف جي 1 سيكلن كيناز يعتمد على سيكلين دي4،6، معارضة فكرة أن نشاطه يزداد تدريجيًا. لا يزال التفاعل آر بي أحادي الفسفور المعتمد على سيكلين دي يتفاعل مع عوامل النسخ إي 2 إف بطريقة تمنع نسخ الإنزيمات التي تدفع انتقال جي 1 على إس. بدلاً من ذلك، يزداد نشاط النسخ المعتمد على إي 2 إف عندما يزيد نشاط سي دي كي2 ويزيد فرط الفسفوريلات آر بي في نهاية جي1. قد لا يكون أر بي هو الهدف الوحيد لـ سيكلين دي لتعزيز تكاثر الخلايا وتطورها خلال دورة الخلية. سيكلن دي- كب سيكلن كيناز يعتمد على سيكلين 4,6، معقد، من خلال الفسفرة وتعطيل الإنزيمات الأيضية، يؤثر أيضًا على بقاء الخلية. من خلال التحليل الدقيق لمختلف اللدائن آر بي دوكينج، تم تحديد عزر تسلسل اللولب الإجماعي، والذي يمكن استخدامه لتحديد الركائز غير القانونية المحتملة التي يمكن أن يستخدمها سيكلين سيكلن كيناز يعتمد على سيكلين دي 4,6 لتعزيز الانتشار.
الالتحام فيفسفور آر بي
تؤثر طفرات الإرساء المستندة إلى آر إس إل وإل في سي في إي بشكل عام على مجمعات سيكلين سي دي كي. تؤدي طفرات مخلفات آر بي الرئيسية التي لوحظت سابقًا على أنها ضرورية لتفاعلات إرساء سي دي كي المعقدة إلى انخفاض نشاط كيناز الكلي تجاه آر بي. انشقاق تجليد إل في سي في إي في مجال الجيب آر بي، والذي ثبت أنه يتفاعل مع بروتينات مثل سيكلين دي والبروتين الفيروسي، لديه انخفاض طفيف بمقدار 1.7 أضعاف في الفسفرة بواسطة سيكلن دي- كب سيكلن كيناز يعتمد على سيكلين كي4،6 عند إزالته. وبالمثل، عندما تتم إزالة الزخم آر في إل، الذي يظهر أنه يتفاعل مع سيكلوس سي، فإن نشاط سيكلن دي- كب سيكلن كيناز يعتمد على سيكلين4،6 ينخفض بمقدار 4.1 أضعاف. وبالتالي، فإن مواقع إرساء آر في إل وإل في سي في إي لها تفاعلات مع سيكلن دي- كب سيكلن كيناز يعتمد على سيكلين4،6 مثلما تفعل مع سيكلانات أخرى، وإزالتها لها تأثير متواضع في تقدم جي 1.
تستهدف مجمعات دي- كب سيكلن كيناز يعتمد على سيكلين 4,6 آلر بي للفسفرة من خلال إرساء حلزون سي الدائم. عندما يتم اقتطاع 37 بقايا من الأحماض الأمينية الأخيرة، فقد ثبت سابقًا أن مستويات الفسفرة آر بي يتم تقليلها ويتم تحفيز جي1. وقد أظهرت الاختبارات الحركية أنه مع نفس الاقتطاع، فإن تقليل الفسفرة آر بي بواسطة سيكلين1 سي دي كي4،6 هو 20 ضعفًا وزاد ثابت ميكايليس مينتين (كيلومتر) بشكل ملحوظ. تتأثر فسفرة آر بي بواسطة سيكلين سي دي كي أ 2 وسيكلين سي دي كي بي1 وسيكلين سي دي كي إي2.
يحتوي الطرف سي على امتداد لـ 21 حمضًا أمينيًا مع ميل ألفا حلزون. كما أن حذف هذا الحلزون أو تعطيله عن طريق استبدال بقايا البرولين يظهر أيضًا انخفاضًا كبيرًا في فسفرة آر بي. يعتبر توجيه البقايا، إلى جانب خصائص القاعدة الحمضية والأقطاب القطبية أمرًا بالغ الأهمية لرسو السفن. وهكذا، تتفاعل جميع مواقع إرساء إل في سي في إي وآر في إلواللولب مع أجزاء مختلفة من سيكلين دي، ولكن تعطيل أي اثنين من الآليات الثلاثة يمكن أن يعطل فسفرة آر بي في المختبر. يعمل حلزون اللولب، وربما الأكثر أهمية، كمتطلب هيكلي. إنه يجعل التطور أكثر صعوبة، مما يؤدي إلى مجمع سيكلين دي -سي دي كي4 / 6 إلى الحصول على عدد صغير نسبيًا من الركائز نسبة إلى مجمعات سيكلينن سي دي كي الأخرى. يساهم هذا في نهاية المطاف في الفسفرة الكافية لهدف رئيسي في المكتب الإقليمي.
جميع مجمعات سيكلين دي- كب سيكلن كيناز يعتمد على سيكلين4،6 الستة (سيكلين دي1 / دي2 / دي3 مع دي- كب سيكلن كيناز يعتمد على سيكلين4 / 6) تستهدف آر بي للفسفرة من خلال الإرساء القائم على اللولب. التصحيح اللولبي المسعور α 1 الذي تمتلكه كل سيكلين دي غير مسؤول عن التعرف على اللولب سي الطرفي. بدلاً من ذلك، يتعرف على تسلسلات آر في إل الخطية، بما في ذلك تلك الموجودة على آر بي. من خلال التجارب مع سيكلين المنقى دي- كب سيكلن كيناز يعتمد على سيكلين2 ، تم التوصل إلى أن موقع إرساء اللولب من المحتمل أن يقع على سيكلين دي بدلاً من دي- كب سيكلن كيناز يعتمد على سيكلين 4,6. نتيجة لذلك، من المحتمل أن تتعرف منطقة أخرى على سيكلين دي على اللولب آر بي سي الدائم.
منذ تثبيط آر بي إس سي - الحلزون الطرفي يربط بشكل حصري دي- كب سيكلن كيناز يعتمد على سيكلين 4,6 وليس معقدات سيكلين السي دي كي الأخرى التي تعتمد على دورة الخلية، من خلال التجارب التي تغير هذا اللولب في خلايا هيميك، لقد ثبت بشكل قاطع أن تفاعل سيكلين دي آر بي هو حاسمة في الأدوار التالية (1) تعزيز انتقال جي1 / إس (2) السماح لتفكك آر بي من الكروماتين، و (3) تنشيط إي2إف1.
اللائحة
في الفقاريات
يتم تنظيم سيكلين دي من خلال مسار المصب لمستقبلات ميتوجين عبر كيناز ماب والراس ومسارات بي كاتنين -تي سي إف / إل إي إف وفوسفوينتيد 3 كيناز . يعمل ماب كيناز إيربك على تنشيط عوامل النسخ النهائية ميس وإي بي-1 وفسفور التي بدورها تنشط نسخ جينات سي دي كي4 وسي دي كي6 وسيكلين دي، وتزيد من التكوين الحيوي للريبوسوم. عائلة رو جي تي فاس، كيناز مرتبط بالتكامل وكيناز الالتصاق البؤري (غف إي كي) ينشط جين سيكلين دي استجابة لـ إنتغرين.
بي27 كيب1 و بي21 سيب1 هي مثبطات كيناز تعتمد على سيكلين (سي كي آي إس) التي تنظم بشكل سلبي سي دي كي إس. ومع ذلك هم أيضا مروجين لمجمع سيكلين دي-سي دي كي4 / 6. بدون بي27 و بي21 ، يتم تقليل مستويات سيكلين دي ولا يتم تكوين المركب عند مستويات قابلة للكشف.
في حقيقيات النوى، يؤدي التعبير الزائد لعامل بدء الترجمة إي 4 (إي آي إف 4 إي) إلى زيادة مستوى بروتين سيكلين دي وزيادة كمية سيكلين دي إم رنا خارج النواة. وذلك لأن إي آي إف 4 إي يعزز تصدير إم رنا إس من سيكلين دي خارج النواة.
تثبيط سيكلين دي عن طريق التعطيل أو التحلل يؤدي إلى خروج دورة الخلية والتمايز. يتم تشغيل تثبيط السيكلين دي عدة تعتمد على السيكلين بروتين كيناز المانع (سي كي آي إس) مثل إنك4 الأسرة (مثل بي14 ، بي15 ، بي16 ، بي18). يتم تنشيط بروتينات إنك4 استجابةً للإجهاد المفرط للتكاثر الذي يمنع تكاثر الخلايا بسبب الإفراط في التعبير عن راس وميك. وبالتالي، يرتبط إنك4 بـ سي دي كي إس المعتمدة على سيكلين دي ويعطل المجمع بأكمله. يسبب الجليكوجين سينثاز كيناز ثلاثة بيتا، جي إس كي 3β ، تدهور سيكلين دي عن طريق الفسفرة المثبطة على ثريونين 286 من بروتين سيكلين دي. يتم التحكم في جي إس كي3β بشكل سلبي من خلال مسار بي آي3 كي في شكل فسفرة، وهي إحدى الطرق العديدة التي تنظم بها عوامل النمو سيكلين دي. النواة وتجميعها مع سي دي كي4 وسي دي كي6.
وقد ثبت أن تثبيط سيكلين دي (سيكلين دي1 و 2، على وجه الخصوص) يمكن أن ينتج عن تحريض بروتين واف1 /سي آي بي / بي21 بواسطة بي دي تي. من خلال تثبيط سيكلين دي، يمنع هذا الاستقراء أيضًا سي دي كي2 و 6. كل هذه العمليات مجتمعة تؤدي إلى القبض على الخلية في مرحلة جي0 / جي1.
هناك طريقتان يؤثر فيه تلف الحمض النووي على سي دي كي إس. بعد تلف الحمض النووي، يتحلل سيكلين (سيكلين1) بسرعة وعابرة بواسطة البروتوسوم. يتسبب هذا التدهور في إطلاق بي21 من مجمعات سي دي كي4 ، والتي تعمل على تعطيل سي دي كي2 بطريقة مستقلة عن بي53. الطريقة الأخرى التي يستهدف بها تلف الحمض النووي سي دي كيs هي الحث المعتمد على بي53 لـ بي21 ، مما يمنع مركبسيكلين إي-سي دي كي2. في الخلايا السليمة، يتحلل بي53 من النوع البري بسرعة بواسطة البروتوسوم. ومع ذلك، فإن تلف الحمض النووي يجعله يتراكم بجعله أكثر استقرارًا.
في الخميرة
التبسيط في الخميرة هو أن جميع السيكلانات ترتبط بنفس الوحدة الفرعية سي دي سي، سي دي سي28. يتم التحكم في السيكلنز في الخميرة عن طريق التعبير، والتثبيط عن طريق سي كي آي إس مثل فار1 ، والتحلل عن طريق التحلل البروتيني بوساطة بروتين يوبيوتيكين.
دور في السرطان
بالنظر إلى أن العديد من السرطانات البشرية تحدث استجابة للأخطاء في تنظيم دورة الخلية وفي المسارات الخلوية التي تعتمد على عامل النمو، فإن إشراك سيكلين دي في التحكم في دورة الخلية وإشارة عامل النمو يجعله ممكنًا في السرطان. في الخلايا الطبيعية، يؤدي الإفراط في إنتاج سيكلين دي إلى تقصير مدة المرحلة جي1 فقط، وبالنظر إلى أهمية سيكلين دي في إشارات عامل النمو، يمكن أن تكون العيوب في تنظيمه مسؤولة عن غياب تنظيم النمو في الخلايا السرطانية. يؤثر الإنتاج غير المنضبط لـ سيكلين دي على كميات مركب سيكلين دي-سي دي كي4 التي يتم تكوينها، والتي يمكن أن تدفع الخلية عبر نقطة تفتيش جي0 على إس، حتى عندما لا تكون عوامل النمو موجودة.
الدليل على أن سيكلين دي1 مطلوب لتكوين الورم يشمل اكتشاف أن تعطيل سيكلين دي1 عن طريق مضاد للحساسية أو حذف الجين يقلل من أورام الثدي ونمو ورم معدي معوي في الجسم الحي. إن التعبير الزائد عن سيكلين دي1 كافٍ لتحريض تكوين الأورام الثديية، يعزى إلى تحريض تكاثر الخلايا، وزيادة بقاء الخلية، تحريض عدم استقرار الصبغيات، تقييد البلعمة الذاتية والوظائف غير القانونية المحتملة.
يحدث الإفراط في التعبير نتيجة لتضخيم الجين أو عامل النمو أو التعبير الناجم عن الجين بواسطة إس آر سي، آر أ إس، إي ى ر بي بي2 ، ستات3 ، ستات5 ، ضعف البروتين، أو نقل الكروموسومات. تضخيم الجينات مسؤول عن الإفراط في إنتاج بروتين سيكلن د في سرطان المثانة وسرطان المريء وغيرها.
في حالات الساركوما وسرطانات القولون والمستقيم والأورام الميلانينية، لوحظ الإفراط في إنتاج سيكلين دي، ولكن بدون تضخيم المنطقة الصبغية التي تشفرها (الكروموسوم صيغة 11 13، الجين الورمي المزعوم بي راد1 ، الذي تم تحديده كحدث نقل في حالة خلايا الوشاح سرطان الغدد الليمفاوية). في أورام الغدة الجار درقية، يحدث فرط إنتاج سيكلين دي بسبب نقل الكروموسومات، مما يضع التعبير عن سيكلين دي (بشكل أكثر تحديدًا، سيكلين دي1) تحت مروج غير مناسب، مما يؤدي إلى الإفراط في التعبير. في هذه الحالة، تم نقل جين سيكلين دي إلى جين هرمون الغدة الجار درقية، وتسبب هذا الحدث في مستويات غير طبيعية من سيكلين دي. لوحظت نفس آليات الإفراط في التعبير عن سيكلين دي في بعض أورام خلايا بائية المنتجة للأجسام المضادة. وبالمثل، لوحظ الإفراط في التعبير عن بروتين سيكلن دي بسبب الانتقال الجيني في سرطان الثدي البشري.
بالإضافة إلى ذلك، يتم تعزيز تطور السرطان أيضًا بحقيقة أن بروتين الورم المثبط لورم الأرومة الشبكية (آر بي)، أحد الركائز الرئيسية لمركب سيكلين سي دي كي 4/6، يتغير بشكل متكرر في الأورام البشرية. في شكله النشط، يمنع آر بي عبور نقطة تفتيش جي 1 عن طريق منع نسخ الجينات المسؤولة عن التقدم في دورة الخلية. سيكلين دي على سي دي كي 4 مركب فسفوريلات آر بي، والذي يعطله ويسمح للخلية بالمرور عبر نقطة التفتيش. في حالة التعطيل غير الطبيعي للرأس، في الخلايا السرطانية، يتم فقدان منظم مهم لتقدم دورة الخلية. عندما يتحور آر بي، تكون مستويات سيكلين دي وبي 16 إنك 4 طبيعية.
منظم آخر للمرور عبر نقطة تقييد جي1 هو مثبط سي دي كي بي16 ، الذي يتم ترميزه بواسطة جين إنك4. وظائف بي16 في تعطيل مركب سيكلين سي دي كي 4. وبالتالي، فإن حظر نسخ جين إنك4 سيزيد من نشاط سيكلين دي سي دي كي4 ، مما يؤدي بدوره إلى تعطيل غير طبيعي لـ آر بي. من ناحية أخرى، في حالة سيكلين دي في الخلايا السرطانية (أو فقدان بي16إنك 4) يتم الاحتفاظ آر بي من النوع البري. نظرًا لأهمية مسار بي16إنك / سيكلين دي / سي دي كي4 أو 6 / آر بي في إشارات عامل النمو، يمكن أن تؤدي الطفرات في أي من اللاعبين المعنيين إلى الإصابة بالسرطان.
النمط الظاهري المسوخ
تشير الدراسات التي أجريت مع المسوخ إلى أن السيكلانات هي منظمات إيجابية لدخول دورة الخلية. في الخميرة، يؤدي التعبير عن أي من سيكلانات جي1 الثلاثة إلى دخول دورة الخلية. نظرًا لأن تقدم دورة الخلية مرتبط بحجم الخلية، تظهر الطفرات في سيكلين دي ومتجانساتها تأخيرًا في دخول دورة الخلية وبالتالي، فإن الخلايا ذات المتغيرات في سيكلين دي لديها أكبر من حجم الخلية الطبيعي في انقسام الخلية.
بي27 - / - خروج المغلوب النمط الظاهري تظهر الإفراط في خلايا لأن السيكلين لا تحول دون دي بعد الآن، في حين بي27 - / - والسيكلين دي - / - بالضربة القاضية تتطور بشكل طبيعي.
انظر أيضا
- CDK
- سيكلنس
- دورة الخلية
المراجع
- ببب: 2W96; Day PJ، Cleasby A، Tickle IJ، O'Reilly M، Coyle JE، Holding FP، وآخرون (مارس 2009). "Crystal structure of human CDK4 in complex with a D-type cyclin". Proceedings of the National Academy of Sciences of the United States of America. ج. 106 ع. 11: 4166–70. DOI:10.1073/pnas.0809645106. PMC:2657441. PMID:19237565.
روابط خارجية
- Cyclin+D في المكتبة الوطنية الأمريكية للطب نظام فهرسة المواضيع الطبية (MeSH).
قالب:بروتينات الخلية
 بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية