اندماج الحويصلة
اندماج الحويصلة هو اندماج حويصلة مع حويصلات أخرى أو جزء من غشاء الخلية. في الحالة الثانية، هي المرحلة النهائية للإفراز من الحويصلات الإفرازية، حيث تُطرد محتوياتها من الخلية من خلال الإخراج الخلوي. يمكن أن تندمج الحويصلات أيضاً مع مقصورات الخلايا المستهدفة الأخرى، مثل الليسوسوم. يحدث الإخراج الخلوي عندما تلتحم الحويصلات الإفرازية بشكل عابر وتندمج عند قاعدة الهياكل ذات شكل الكوب في غشاء بلازما الخلية المسمى بوروسوم، وهي آلية إفرازية غالبة في الخلايا. قد يعتمد اندماج الحويصلة على بروتينات سنير في وجود زيادة تركيز الكالسيوم داخل الخلايا (+Ca2).
محفزات
تعمل المحفزات التي تؤدي إلى اندماج الحويصلة بزيادة +Ca2 داخل الخلايا.
- الحويصلات المشبكية تُلزِم اندماج الحويصلة بالحدوث عن طريق نبض عصبي يصل إلى المشبك، مما يؤدي إلى تنشيط قنوات الكالسيوم المعتمدة على الجهد والتي تسبب تدفق +Ca2 إلى الخلية.
- في جهاز الغدد الصماء، تُطلق العديد من الهرمونات بوساطة هرموناتها المُفرِزة التي ترتبط بمستقبلات مقترنة بالبروتين G المرتبطة بالوحدة الفرعية ألفا Gq، مما يؤدي إلى تنشيط مسار IP3/DAG لزيادة +Ca2. ومن أمثلة هذه الآلية ما يلي:
- هرمون إفراز الغدد التناسلية.[1]
- هرمون إفراز الثيروتروبين.[1]
- هرمون مطلق لهرمون النمو[1] (المسار الثانوي-المسار الرئيسي هو المسار المعتمد على cAMP).[2]
أنظمة نموذجية
دُرِست الأنظمة النموذجية التي تتكون من فوسفوليبيد واحد أو خليط من قبل الكيميائيين الفيزيائيين.
يوجد الكارديوليبين بشكل رئيسي في أغشية الميتوكوندريون، وتلعب أيونات الكالسيوم دوراً مهماً في العمليات التنفسية بوساطة الميتوكوندريون. افتُرِضَت القوى المتضَمة لشرح[3] هذه العملية من حيث تكوين نواة لتكتل الكيانات فوق الجزيئية الأصغر أو تغيرات الطور في بنية الأغشية الحيوية.[4]
آليات
اندماج الشق المشبكي
يجب أن تكون الحويصلة على بعد بضعة نانومترات من الغشاء المستهدف حتى تبدأ عملية الاندماج في الحويصلة المشبكية. يسمح هذا التقارب لغشاء الخلية والحويصلة بتبادل الدهون التي تتوسطها بروتينات معينة تزيل الماء الذي يأتي بين الارتباط المكون. بمجرد أن تصبح الحويصلة في موضعها، يجب أن تنتظر حتى يدخل + Ca2 الخلية عن طريق انتشار جهد الفعل إلى الغشاء قبل المشبكي.[5] يرتبط +Ca2 ببروتينات معينة، أحدها هو سينابتوتاجمين، ذلك في الخلايا العصبية التي تؤدي إلى الاندماج الكامل للحويصلة مع الغشاء المستهدف.[6]
يُعتقد أيضاً أن بروتينات سنير تساعد في التوسط في الغشاء المستهدف للحويصلة.[7]
بروتين سنير وتشكيل المسام
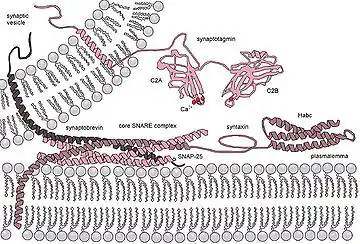
يُحتمل أن يؤدي تجميع بروتينات سنير في التركيبات "العابرة" إلى ربط الطبقات الثنائية الدهنية المتعارضة للأغشية التي تنتمي إلى الخلية والحبيبات الإفرازية، مما يجعلها قريبة وتحفز اندماجها. يؤدي تدفق الكالسيوم إلى الخلية إلى اكتمال تفاعل التجميع، والذي يكتمل بوساطة التفاعل بين مستشعر الكالسيوم المفترض، سينابتوتاجمين، مع الدهون الغشائية و/أو تركيب سنير المجمع جزئياً.
تتضمن إحدى الفرضيات جزيء كومبلكسن داخل مجمع سنير وتفاعله مع جزيء سينابتوتاجمين.[9] المعروفة باسم فرضية "المشبك"، وجود المركب عادةً ما يمنع اندماج الحويصلة في غشاء الخلية. ومع ذلك، فإن ربط أيونات الكالسيوم ب سينابتوتاجمين يؤدي إلى إطلاق المركب أو تعطيله، بحيث تكون الحويصلة حرة في الاندماج.[10]
وفقاً لفرضية "السحاب/الزمام المغلق"، يبدأ التجميع المعقد عند الأجزاء الطرفية N من أنماط سنير ويتقدم نحو طرف C التي تثبت البروتينات المتفاعلة في الأغشية. يستمر تكوين مركب سنير "العابر" من خلال مركب وسيط يتكون من سناب-25 وسينتاكسين-1، والذي يستوعب لاحقاً سينابتوبريفين-2 (تشارك الأنماط المتماثلة سينتاكسين وسينابتوبريفين المقتبسة في إطلاق الوسيط العصبي العصبوي).
استناداً إلى استقرار مجمع سيس سنيرالناتج، فقد افتُرِضَ أن الطاقة المنبعثة أثناء عملية التجميع تعمل كوسيلة للتغلب على قوى التنافر بين الأغشية. هناك العديد من النماذج التي تقترح تفسيراً لخطوة لاحقة - تشكيل مسام الساق والانصهار، لكن الطبيعة الدقيقة لهذه العمليات لا تزال موضع نقاش. اثنان من أبرز النماذج في تكوين مسام الانصهار هما نظريات مسام الانصهار المبطنة بالدهون والبروتين.[11]
نظرية مسام الاندماج المبطنة بالدهون
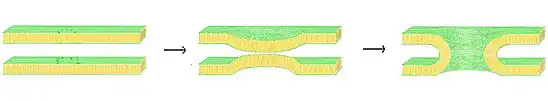
أحد النماذج الممكنة لتشكيل مسام الاندماج هو نظرية المسام المبطنة بالدهون. في هذا النموذج، بمجرد أن تكون الأغشية قريبة بما فيه الكفاية عبر آلية "السحاب/الزمام المغلق" لمركب سنير، يحدث اندماج الغشاء تلقائياً. لقد ثبت أنه عندما يُحضر الغشاءين ضمن مسافة حرجة، فمن الممكن أن تندمج مجموعات رؤوس الدهون المحبة للماء لغشاء واحد مع الغشاء المقابل.[12] في نموذج مسام الاندماج المبطنة بالدهون، يعمل مركب سنير كسقالة، يسحب الغشاء، مما يتسبب في تجعد كلا الغشاءين حتى يصلا إلى مسافة الاندماج الحرجة. عندما يبدأ الغشاءان في الاندماج،يُنتج ساق مبطن بالدهون، يتوسع شعاعياً للخارج مع استمرار الاندماج.
حيث أن المسام المبطنة بالدهون ممكنة وتحقق جميع الخصائص نفسها التي لوحظت في تكوين المسام المبكر، لا توجد بيانات كافية لإثبات أنها الطريقة الوحيدة للتكوين.[13] لا توجد حالياً آلية مقترحة للتنظيم بين الخلايا لتذبذب المسام المبطنة بالدهون، وستواجه وقتاً أكثر صعوبة في إنتاج تأثيرات مثل "القبلة والجري" عند مقارنتها بنظيراتها المبطنة بالبروتين. تعتمد فعالية المسام المبطنة بالدهون أيضاً بشكل كبير على تكوين كلا الغشاءين، ويمكن أن يختلف نجاحها أو فشلها بشكل كبير مع التغيرات في المرونة والصلابة.[13]
نظرية مسام الانصهار المبطنة بالبروتين
نموذج آخر محتمل لتشكيل مسام الاندماج هو نظرية المسام المبطنة بالبروتين. في هذا النموذج، بعد تنشيط سينابتوتاجمين بواسطة الكالسيوم، تتجمع العديد من مجمعات سنير معاً لتشكيل بنية حلقية، حيث يشكل سينابتوبريفين المسام في غشاء الحويصلة ويشكل سينتاكسين المسام في غشاء الخلية.[14] مع توسع المسام الأولية، فإنها تشتمل على دهون من كلا الطبقتين، مما يؤدي في النهاية إلى اندماج كامل بين الغشاءين. مركب سنير له دور أكثر نشاطاً في نظرية المسام المبطنة بالبروتين. نظراً لأن المسام تتكون في البداية بالكامل من بروتينات سنير، فإن المسام قادرة بسهولة على الخضوع للتنظيم بين الخلايا، مما يجعل التذبذب وآليات "القبلة والهروب" سهلة الحدوث.[9]
تلبي المسام المبطنة بالبروتين تماماً جميع المتطلبات المرصودة لمسام الاندماج المبكر، وبينما تدعم بعض البيانات هذه النظرية،[14] لا توجد بيانات كافية لاعتمادها كطريقة أساسية للاندماج. تتطلب المسام المبطنة بالبروتين خمس نسخ على الأقل من مركب سنير بينما لوحظ الاندماج مع عدد قليل من نسختين.[14]
تظل وظيفة معقد سنير دون تغيير إلى حد كبير، في كلتا النظريتين، ومجمع سنير بأكمله ضروري لبدء الاندماج. ومع ذلك، فقد ثبت أن سينتاكسين في المختبر في حد ذاته يكفي لدفع الاندماج التلقائي المستقل للكالسيوم للحويصلات المشبكية التي تحتوي على سنير-v.[15] هذا يشير إلى أن سينابتوتاجمين الإخراج الخلوي العصبي المعتمد على +Ca2 هو منظم مزدوج، في غياب أيونات +Ca2 لتثبيط ديناميكيات سنير، بينما في وجود أيونات +Ca2 لتكون بمثابة ناهض في عملية اندماج الغشاء.
فرضية القبلة والهرب
اقترح بعض علماء الكيمياء العصبية أن الحويصلات المشبكية في بعض الأحيان قد لا تندمج تماماً مع الأغشية قبل المشبكية في إطلاق الناقل العصبي في الشق المشبكي. يكمن الجدل في ما إذا كان الإدخال الخلوي يحدث دائماً في إصلاح الحويصلة بعد إطلاق الناقل العصبي أم لا. آلية أخرى مقترحة لإطلاق محتويات الحويصلة في السائل خارج الخلية تسمى الاندماج بالقبلة والجري.
توجد بعض الدلائل على أن الحويصلات قد تشكل فقط مسام صغيرة في الغشاء قبل المشبكي مما يسمح بإطلاق المحتويات عن طريق الانتشار القياسي لفترة قصيرة قبل التراجع مرة أخرى إلى الخلية قبل المشبكية. قد تكون هذه الآلية طريقة للتغلب على الإدخال الخلوي بوساطة الكلثرين. يقترح أيضاً أن الحويصلة لا تحتاج إلى العودة إلى الإندوزوم لإعادة ملئها، على الرغم من أنه ليس مفهوماً تماماً من خلال الآلية التي ستعيد تعبئتها. هذا لا يستبعد اندماج الحويصلة الكاملة، ولكنه ينص فقط على أن كلتا الآليتين قد تعملان في الشقوق المشبكية.
ثبت أن "القبلة والجري" تحدث في خلايا الغدد الصماء، على الرغم من أنها لم تشاهد مباشرة في الفجوات المشبكية.[16]
انظر أيضاً
- الجسيمات الشحمية المستخدَمة كنماذج للخلايا الاصطناعية في دراسات اندماج الأغشية.
المراجع
- Page 237 in: Costanzo, Linda S. (2007). Physiology. Hagerstwon, MD: Lippincott Williams & Wilkins. ISBN:978-0-7817-7311-9. مؤرشف من الأصل في 2022-07-25.
- Walter F., PhD. Boron (2003). Medical Physiology: A Cellular And Molecular Approaoch. Elsevier/Saunders. ص. 1300. ISBN:978-1-4160-2328-9.
- Papahadjopoulos، Demetrios (1990). "Molecular mechanisms of calcium-induced membrane fusion". Journal of Bioenergetics and Biomembranes. ج. 22 ع. 2: 157–179. DOI:10.1007/BF00762944. PMID:2139437. S2CID:1465571.
- sciencedirect نسخة محفوظة 2021-12-03 على موقع واي باك مشين.
- Pigino، Gustavo؛ Morfini، Gerardo؛ Brady، Scott (2006). "Chapter 9: Intracellular Trafficking". في Siegal، George J.؛ Albers، R. Wayne؛ Brady، Scott T.؛ وآخرون (المحررون). Basic Neurochemistry: Molecular, Cellular and Medical Aspects (Textbook) (ط. 7th). Burlington, MA: Elsevier Academic Press. ص. 143. ISBN:978-0-12-088397-4.
- Pigino et al. p 158
- Pigino et al. p.143
- Georgiev، Danko D .؛ James F . Glazebrook (2007). "Subneuronal processing of information by solitary waves and stochastic processes". في Lyshevski، Sergey Edward (المحرر). Nano and Molecular Electronics Handbook. Nano and Microengineering Series. CRC Press. ص. 17–1–17–41. DOI:10.1201/9781315221670-17. ISBN:978-0-8493-8528-5. S2CID:199021983.
- Kümmel، D.؛ Krishnakumar، S. S.؛ Radoff، D. T.؛ Li، F.؛ Giraudo، C. G.؛ Pincet، F.؛ Rothman، J. E.؛ Reinisch، K. M. (2011). "Complexin cross-links prefusion SNAREs into a zigzag array". Nature Structural & Molecular Biology. ج. 18 ع. 8: 927–933. DOI:10.1038/nsmb.2101. PMC:3410656. PMID:21785414.
- Richmond، Janet. "Synapse Function". مؤرشف من الأصل في 2022-12-24.
- Jackson، Meyer B.؛ Chapman، Edwin R. (2006). "Fusion Pores and Fusion Machines in Ca2+-Triggered Exocytosis". Annual Review of Biophysics and Biomolecular Structure. ج. 35 ع. 1: 135–160. DOI:10.1146/annurev.biophys.35.040405.101958. PMID:16689631.
- Marrink، Siewert J.؛ Mark، Alan E. (1 سبتمبر 2003). "The Mechanism of Vesicle Fusion as Revealed by Molecular Dynamics Simulations" (PDF). Journal of the American Chemical Society. ج. 125 ع. 37: 11144–11145. DOI:10.1021/ja036138+. ISSN:0002-7863. PMID:16220905. مؤرشف من الأصل (PDF) في 2023-01-28.
- Nanavati، C؛ Markin، V S؛ Oberhauser، A F؛ Fernandez، J M (1 أكتوبر 1992). "The exocytotic fusion pore modeled as a lipidic pore". Biophysical Journal. ج. 63 ع. 4: 1118–1132. Bibcode:1992BpJ....63.1118N. DOI:10.1016/s0006-3495(92)81679-x. ISSN:0006-3495. PMC:1262250. PMID:1420930.
- Chang، Che-Wei؛ Hui، Enfu؛ Bai، Jihong؛ Bruns، Dieter؛ Chapman، Edwin R.؛ Jackson، Meyer B. (8 أبريل 2015). "A Structural Role for the Synaptobrevin 2 Transmembrane Domain in Dense-Core Vesicle Fusion Pores". The Journal of Neuroscience. ج. 35 ع. 14: 5772–5780. DOI:10.1523/JNEUROSCI.3983-14.2015. ISSN:0270-6474. PMC:4388931. PMID:25855187.
- Woodbury DJ, Rognlien K (2000). "The t-SNARE syntaxin is sufficient for spontaneous fusion of synaptic vesivles to planar membranes" (PDF). Cell Biology International. ج. 24 ع. 11: 809–818. DOI:10.1006/cbir.2000.0631. PMID:11067766. S2CID:37732173. مؤرشف من الأصل (PDF) في 2011-07-19. اطلع عليه بتاريخ 2009-05-31.
- Piginio et al. pp. 161-162
 بوابة طب
بوابة طب