المركب التنفسي
المركب التنفسي I ، EC 7.1.1.2 (المعروف أيضًا باسم NADH: ubiquinone oxidoreductase ، Type I NADH نازعة الهيدروجين و مجمع الميتوكوندريا I) هو أول مجمع بروتين كبير لسلاسل الجهاز التنفسي للعديد من الكائنات الحية من البكتيريا إلى البشر. يحفز نقل الإلكترونات من NADH إلى الإنزيم المساعد Q10 (CoQ10) وينقل البروتونات عبر غشاء الميتوكوندريا الداخلي في حقيقيات النوى أو غشاء البلازما للبكتيريا.
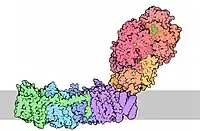
هذا الإنزيم ضروري للتشغيل الطبيعي للخلايا والطفرات في وحداتها الفرعية التي تؤدي إلى مجموعة واسعة من الاضطرابات العصبية والعضلية الموروثة والتمثيل الغذائي. العيوب في هذا الإنزيم مسؤولة عن تطوير العديد من العمليات المرضية مثل تلف نقص التروية / إعادة ضخ الدم (السكتة الدماغية واحتشاء القلب) ، ومرض باركنسون وغيرها.
تشير الأبحاث الحديثة إلى أن المركب I هو مصدر قوي لأنواع الأكسجين التفاعلية. يمكن أن ينتج المركب الأول أكسيدًا فائقًا (بالإضافة إلى بيروكسيد الهيدروجين) ، من خلال مسارين مختلفين على الأقل. أثناء نقل الإلكترون الأمامي ، يتم إنتاج كميات صغيرة جدًا من الأكسيد الفائق (ربما أقل من 0.1٪ من إجمالي تدفق الإلكترون).
أثناء النقل الإلكتروني العكسي ، قد يكون المركب الأول هو أهم موقع لإنتاج الأكسيد الفائق داخل الميتوكوندريا ، حيث يتم تحويل حوالي 3-4٪ من الإلكترونات إلى تكوين الأكسيد الفائق.[1] نقل الإلكترون العكسي ، وهي العملية التي تمر بها الإلكترونات من تجمع ubiquinol المخفض (يتم توفيره من خلال ديهيدروجيناز السكسينات ، ديهيدروجيناز الجلسرين 3-فوسفات ، إلكترون فلافوبروتين أو ديهيدروروتيت ديهيدروجينيز في الميتوكوندريا الثديية) عبر المركب I لتقليل NAD + إلى NADH ، الجهد الكهربائي المحتمل لغشاء الميتوكوندريا الداخلية. على الرغم من أنه لا يُعرف على وجه الدقة تحت الظروف المرضية التي يمكن أن يحدث فيها انتقال الإلكترون العكسي في الجسم الحي ، إلا أن التجارب في المختبر تشير إلى أن هذه العملية يمكن أن تكون مصدرًا قويًا جدًا للأكسيد عندما تكون تركيزات السكسينات عالية وتركيزات أكسالات أو مالات منخفضة.[2] يمكن أن يحدث هذا أثناء نقص تروية الأنسجة ، عندما يتم حظر توصيل الأكسجين.
الآلية الشاملة
تحدث جميع تفاعلات الأكسدة والاختزال في المجال المحبة للماء للمركب الأول. ويرتبط NADH في البداية بالمركب الأول ، وينقل إلكترونين إلى مجموعة الأطراف الاصطناعية من الفلافين أحادي النوكليوتيد (FMN) من الإنزيم ، مما يخلق FMNH2. إن متقبل الإلكترون - حلقة isoalloxazine - من FMN مطابق لتلك الموجودة في FAD. يتم بعد ذلك نقل الإلكترونات عبر FMN عبر سلسلة من مجموعات الكبريت الحديدي (Fe-S) ، وأخيرًا إلى الإنزيم المساعد Q10 (ubiquinone). يغير تدفق الإلكترون هذا حالة الأكسدة للبروتين ، مما يؤدي إلى تغيرات مطابقة للبروتين والتي تغير قيم pK للسلسلة الجانبية المؤينة ، وتتسبب في ضخ أربعة أيونات الهيدروجين من مصفوفة الميتوكوندريا.[3]
آلية التحويل الإلكتروني
المسار المقترح لنقل الإلكترون قبل اختزال ubiquinone هو كما يلي: NADH - FMN - N3 - N1b - N4 - N5 - N6a - N6b - N2 - Q ، حيث Nx هو عبارة عن اتفاقية وضع العلامات على مجموعات كبريت الحديد.[4] تتيح إمكانية الاختزال العالية لمجموعة N2 والقرب النسبي للمجموعات الأخرى في السلسلة نقل الإلكترون الفعال لمسافات طويلة في البروتين (بمعدلات نقل من NADH إلى N2 من مجموعة كبريت الحديد حوالي 100 ميكرومتر).[5][6]
إن ديناميكيات التوازن للمركب I مدفوعة بشكل أساسي بدورة الأكسدة والاختزال الكينوني. في ظروف قوة دافعة عالية للبروتون (وبالتالي ، تجمع مركز ubiquinol) ، يعمل الإنزيم في الاتجاه العكسي. يتأكسد اليوبيكوينول إلى اليوبيكوينون ، وتقلل البروتونات المنبعثة الناتجة من قوة دافع البروتون.
آلية انتقال البروتون
يُقترح حاليًا اقتران نقل البروتون ونقل الإلكترون في المجمع الأول على أنه غير مباشر (تغييرات مطابقة طويلة المدى) على عكس المباشر (وسيطات الأكسدة والاختزال في مضخات الهيدروجين كما هو الحال في مجموعات الهيم من المجمعين الثالث والرابع). تُظهر بنية المنطقة المسعورة للمركب الأول العديد من ناقلات البروتونات المترابطة ميكانيكيًا. المكونات المركزية الثلاثة التي يُعتقد أنها تساهم في حدث التغيير المطابق بعيد المدى هي مجموعة كبريتات الحديد N2 المقترنة بالأس الهيدروجيني ، وتقليل الكينون ، ووحدات اللولب الفرعية للذراع الغشائي. يمكن أن يؤدي نقل التغييرات المطابقة لقيادة ناقلات الغشاء المرتبطة بواسطة `` قضيب توصيل أثناء اختزال ubiquinone إلى اثنين أو ثلاثة من البروتونات الأربعة التي يتم ضخها لكل NADH المؤكسد. يجب ضخ البروتون المتبقي عن طريق الاقتران المباشر في موقع ربط ubiquinone. يقترح أن آليات الاقتران المباشر وغير المباشر مسؤولة عن ضخ البروتونات الأربعة.
 بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة علم الأحياء
بوابة علم الأحياء
المصادر
- Stepanova، Anna؛ Kahl، Anja؛ Konrad، Csaba؛ Ten، Vadim؛ Starkov، Anatoly S.؛ Galkin، Alexander (ديسمبر 2017). "Reverse electron transfer results in a loss of flavin from mitochondrial complex I: Potential mechanism for brain ischemia reperfusion injury". Journal of Cerebral Blood Flow and Metabolism. ج. 37 ع. 12: 3649–3658. DOI:10.1177/0271678X17730242. ISSN:1559-7016. PMC:5718331. PMID:28914132.
- Muller FL، Liu Y، Abdul-Ghani MA، Lustgarten MS، Bhattacharya A، Jang YC، Van Remmen H (يناير 2008). "High rates of superoxide production in skeletal-muscle mitochondria respiring on both complex I- and complex II-linked substrates". The Biochemical Journal. ج. 409 ع. 2: 491–9. DOI:10.1042/BJ20071162. PMID:17916065.
- {{استشهاد بكتاب | عنوان = Principles of Biochemistry, 3rd Edition | chapter = Chapter 18, Mitochondrial ATP synthesis | مؤلف1 = Donald J. Voet | مؤلف2 = Judith G. Voet | مؤلف3 = Charlotte W. Pratt | ناشر = Wiley | سنة = 2008 | isbn = 978-0-470-23396-2 | صفحة =
- Sazanov LA (يونيو 2015). "A giant molecular proton pump: structure and mechanism of respiratory complex I". Nature Reviews. Molecular Cell Biology. ج. 16 ع. 6: 375–88. DOI:10.1038/nrm3997. PMID:25991374.
- Ohnishi، T (1998). "Iron–sulfur clusters/semiquinones in complex I". Biochim. Biophys. Acta. ج. 1364 ع. 2: 186–206. DOI:10.1016/s0005-2728(98)00027-9. PMID:9593887.
- Bridges HR، Bill E، Hirst J (يناير 2012). "Mössbauer spectroscopy on respiratory complex I: the iron-sulfur cluster ensemble in the NADH-reduced enzyme is partially oxidized". Biochemistry. ج. 51 ع. 1: 149–58. DOI:10.1021/bi201644x. PMC:3254188. PMID:22122402.