المجموعة الفردانية E-M2
المجموعة الفردانية E-M2 أو هابلوغروب E-M2، والمعروفة أيضًا باسم E1b1a1-M2، هي مجموعة هابلوغروبية من الحمض النووي لكروموسوم Y البشري.
| النمط الجماعي E-M2 (former E3a / E1b1a) | |
|---|---|
| الوقت المحتمل للنشوء | 39,200 سنة مضت |
| المكان المحتمل للنشوء | غرب إفريقيا أو وسط إفريقيا |
| السلف | E-V38 |
| الطفرة المعرّفة | M2, DYS271/SY81, M291, P1/PN1, P189.1, P293.1 |
ينتشر E-M2 بشكل أساسي داخل إفريقيا جنوب الصحراء الكبرى.
وبشكل أكثر تحديدًا، E-M2 هي الفئة الفرعية السائدة في غرب إفريقيا ووسط إفريقيا وجنوب إفريقيا ومنطقة البحيرات الكبرى الإفريقية؛ كما أنه يحدث بترددات منخفضة إلى متوسطة في شمال إفريقيا، وبترددات منخفضة في الشرق الأوسط.
تحتوي E-M2 على عدة فئات فرعية، ولكن يتم تضمين العديد من هذه المجموعات الفرعية إما في E-L485 أو E-U175.
E-M2 شائع بشكل خاص بين السكان الأصليين الأفارقة الذين يتحدثون لغات النيجر والكونغو، وانتشر إلى جنوب إفريقيا وشرق أفريقيا من خلال توسع البانتو.
الأصول
أُكتشف اثنين من تعدد الأشكال (V38 وV100) بواسطة ترومبيتا في عام 2011 م وقد أعاد تعريف شجرة النشوء والتطور لـ E-V38 بشكل ملحوظ.
وقد دفع هذا المؤلفين إلى الإشارة إلى أن E-V38 ربما نشأ في شرق إفريقيا.
يتبع E-M2 إلى E-V38 والمنتشر في غرب إفريقيا مع سلف مشترك سابق (E-P2)، وربما قد نشأت أيضًا في شرق إفريقيا.
ويتبع E-M329 أيضًا إلى E-V38 والمنتشر في شمال شرق إفريقيا مع سلف مشترك سابق (E-P2)، وربما قد نشأت أيضًا في شرق إفريقيا.
قد يكون SNP E-M180 قد نشأ في اتجاه مجرى النهر في السافانا / الأراضي العشبية الرطبة في جنوب وسط الصحراء الكبرى في شمال إفريقيا ما بين 14000 سنة مضت و 10000 سنة مضت.
ووفقًا لوود في 2005، وروزا في 2007، غيرت هذه الحركات السكانية تنوع الكروموسومات Y الموجود مسبقًا في وسط وجنوب وجنوب شرق إفريقيا، لتحل محل ترددات المجموعات الفردانية السابقة (المجموعات الفردانية A و B-M60) في هذه المناطق بسلالات E1b1a1 المهيمنة الآن.
ومع ذلك، يمكن ملاحظة آثار السكان الأوائل اليوم في هذه المناطق من خلال وجود مجموعات Y DNA الفردانية A1a وA1b وA2 وA3 و B-M60 الشائعة في مجموعات سكانية معينة مثل مبوتي وكويسان.
أشار شرينر في 2018 بالمثل إلى أن المجموعة الفردانية E1b1a-V38 هاجرت عبر الصحراء الخضراء من الشرق إلى الغرب منذ حوالي 19000 عام، حيث ربما نشأت E1b1a1-M2 لاحقًا في غرب إفريقيا أو وسط إفريقيا.
نشر شرينر في 2018 تتبعه أيضًا هذه الهجرة عبر داء الخلايا المنجلية، والتي من المحتمل أنها نشأت خلال فترة الصحراء الخضراء.
الحمض النووي القديم
بوتسوانا
في تاوكومي، في بوتسوانا ، كان هناك فرد يعود تاريخه إلى العصر الحديدي المبكر (1100 سنة مضت)، يحمل المجموعات الفردانية E1b1a1 (E-M2، E-Z1123) و L0d3b1.[1] [2]
في زارو، في بوتسوانا ، كان هناك شخصان يعود تاريخهما إلى العصر الحديدي المبكر (1400 سنة مضت)؛ واحدة تحمل المجموعات الفردانية E1b1a1a1c1a و L3e1a2، والأخرى تحمل المجموعات الفردانية E1b1b1b2b (E-M293، E-CTS10880) وL0k1a2.[1] [2]
جمهورية الكونغو الديمقراطية
في كيندوكي في جمهورية الكونغو الديمقراطية، كان هناك ثلاثة أفراد، يعود تاريخهم إلى فترة ما قبل التاريخ (230 قبل الحاضر، 150 قبل الحاضر، 230 قبل الحاضر)؛ حملت إحداهما المجموعات الفردانية E1b1a1a1d1a2 (E-CTS99، E-CTS99) وL1c3a1b، وحملت الأخرى المجموعة الفردانية E (E-M96، E-PF1620)، والأخيرة حملت المجموعات الفردانية R1b1 (R-P25 1، R-M415) وL0a1b1a1.[1] [2]
مصر
عزم حواس في 2012 م على أن المومياء المصرية القديمة لرجل مجهول مدفون مع رمسيس كانت بسبب العلاقة الوراثية المثبتة وعملية التحنيط التي توحي بالعقاب، مرشحًا جيدًا لابن الفرعون بنتاوير، الذي كان الابن الوحيد الذي ثار ضده أبوه.[3]
وكان من المستحيل تحديد سبب وفاته.[3]
باستخدام تنبؤ مجموعة هابلوغروب الخاصة بـ Whit Athey استنادًا إلى قيم Y- STR، كان من المتوقع أن تشترك كلتا المومياوات في مجموعة هابلوغروب Y الكروموسومية E1b1a1-M2 و50% من مادتهما الوراثية، مما يشير إلى وجود علاقة بين الأب والابن.[3]
أشار جاد في 2021 م إلى أن رمسيس الثالث والرجل المجهول E نتيجته نفس نتيجة بنتاوير، كانا يحملان المجموعة الفردانية E1b1a.[4]
كينيا
في مزرعة ديلورين في ناكورو، كينيا، حمل عالم معادن الحديد في العصر الحديدي المجموعات الفردانية E1b1a1a1a1a/E-M58 وL5b1.[5] [6]
في لامو في جزيرة بات، فازا، في كينيا، كان هناك فرد يعود تاريخه إلى ما بين 1500 م و1700 م، يحمل المجموعتين الفردانيتين E1b1a1a1a2a1a وL3e3a.[7]
في تايتا تافيتا، ماكواسيني في كينيا، كان هناك فرد يعود تاريخه إلى ما بين 1650 م و1950 م، يحمل المجموعتين الفردانيتين E1b1a1a1a2a1a وL4b2a.[7]
في تايتا تافيتا، ماكواسيني في كينيا، كان هناك فرد يعود تاريخه إلى ما بين 1650 م و1950 م، يحمل المجموعتين الفردانيتين E1b1a1a1a2a1a3b1d1c وL1c3b1a.[7]
في تايتا تافيتا، ماكواسيني في كينيا، كان هناك فرد يعود تاريخه إلى ما بين 1650 م و1950 م، يحمل المجموعتين الفردانيتين E1b1a1a1a2a1a وL2a1+143.[7]
في تايتا تافيتا، ماكواسيني في كينيا ، كان هناك فرد يرجع تاريخه إلى ما بين 1667 م و1843 م، يحمل المجموعات الفردانية E1b1a1a1a2a1a3b1d1c وL2a1+143.[7]
في تايتا تافيتا، ماكواسيني في كينيا، كان هناك فرد يرجع تاريخه إلى ما بين 1709 م و1927 م، يحمل المجموعات الفردانية E1b1a1a1a2a1a3a1d~ وL3a2.[7]
تنزانيا
في سونغو منارا في تنزانيا، كان هناك فرد يرجع تاريخه إلى ما بين 1418 م و1450 م، يحمل المجموعتين الفردانيتين E1b1a1~ وL3e2b.[7]
في إقليم ليندي في تنزانيا، كان هناك فرد يرجع تاريخه إلى ما بين 1511 م و1664 م، يحمل المجموعتين الفردانيتين E1b1a1a1a2a1a3a1d~ وL0a1a2.[7]
المكسيك
في موقع دفن مستشفى سان خوسيه دي لوس ناتوراليس الملكي، في مكسيكو سيتي، المكسيك، ثلاثة مستعبدين من غرب إفريقيا من أصول غرب إفريقية وجنوب إفريقية، يعود تاريخهم إلى ما بين 1453 م و1626 م، و1450 م و1620 م، و1436 م، و1472 م.، تم ايجادهم واحدة تحمل المجموعات الفردانية E1b1a1a1c1b/E-M263.2 وL1b2a، وأخرى تحمل المجموعات الفردانية E1b1a1a1d1/E-P278.1/E-M425 و L3d1a1a، والأخيرة تحمل المجموعات الفردانية E1b1a1a1c1a1c/E-CTS8030 و L3e1a1a.[8]
تؤكد أليلات مستضد الكريات البيض البشرية أيضًا أن الأفراد كانوا من أصل إفريقي جنوب الصحراء الكبرى.[9]
البرتغال
في كابيكو دا أموريرا في البرتغال، تم دفن رجل مستعبد من غرب إفريقيا، ربما كان من منطقة سنغامبيا الساحلية في غامبيا أو موريتانيا أو السنغال، وكان يحمل المجموعتين الفردانيتين E1b1a وL3b1a، بين تل ميدنز بين القرن السادس عشر الميلادي والقرن الثامن عشر الميلادي.[10]
سانت هيلينا
في سانت هيلينا، كان 20 إفريقيًا محررًا، [11] [12] يعود تاريخهم إلى القرن التاسع عشر الميلادي، [11] كانوا أيضًا من غرب إفريقيا الوسطى[11] [13] [14] (على سبيل المثال، شعوب البانتو في الجابون وأنغولا) أسلاف.[11]
إحدى الإناث حملت المجموعة الفردانية L1b1a10b.[15]
إحدى الإناث حملت المجموعة الفردانية L2a1f.[15]
إحدى الإناث حملت المجموعة الفردانية L2a1a3c.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a2a1a3b1d و L1c3a.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a1c1a1a و L0a1b2a.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a2a1a3b1a2a2 و L0a1e.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a2a1a3b1 و L2a1f1.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1 و L3.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a2a1a3b1d و L3e1e.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a2a1a3a1d و L3e3b2.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a1c1a1a3 و L3e1a3a.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a2a1a3b1a2a2 و L2b1a.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a2a1a3b1 و L3f1b1a.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a2a1a3b1d1c1a و L3d3a1.[15]
أحد الذكور حمل المجموعات الفردانية B2a1a1a1 و L3e2b1.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a2a1a3b1d1c1a و L2a1f.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a1c1a1a3a1c1 و L3e1d1a.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a2a1a3a1d و L1b1a10.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a1c1a1a3a1c و L2a1f1.[15]
أحد الذكور حمل المجموعات الفردانية E1b1a1a1a1c1a1 و L2b1a.[15]
رجل وامرأة أمريكيان من أصل إفريقي مستعبدان، من موقع دفن شارع أنسون في القرن الثامن عشر الميلادي في تشارلستون، كارولينا الجنوبية، حملوا المجموعة الفردانية L3e1e، شاركت هذه المجموعة الفردانية مع الأفارقة المحررين في سانت هيلينا.[16]
استنادًا إلى أولئك الذين كانوا موجودين بين الأفارقة المستعبدين، فإن نسبة الذكور إلى الإناث تدعم الاستنتاج بوجود تحيز قوي في اختيار الذكور في الفترة الأخيرة من تجارة العبيد عبر المحيط الأطلسي.[11] [17] [18]
ونتيجة لذلك، وبسبب هذه الدراسة التي أجريت على الأفارقة المحررين في سانت هيلينا، من بين دراسات أخرى، تم تقديم رؤى جينية أكبر حول تجارة العبيد عبر المحيط الأطلسي وتأثيراتها على التركيبة السكانية لإفريقيا.[19]
إسبانيا
في غرناطة ، مسلم ( موري ) من خلافة قرطبة،[20] كان من المجموعات الفردانية E1b1a1 و H1+16189،[21] [22] ويقدر تاريخه أيضًا بين 900 م و 1000 م، وموريسكي،[20] الذي كان من المجموعة الفردانية L2e1،[21] [22] وكذلك يقدر تاريخه بين 1500 م و1600 م، وقد وجد أن كلاهما من غرب إفريقيا (أي غامبيا) ومن أصل أيبيري.[20]
الولايات المتحدة الأمريكية
في استراحة أيفري، في تشيسابيك، ديلاوير، كان هناك 3 من أصل 11 فردًا أمريكيين من أصل أفريقي، والذين يرجع تاريخهم إلى ما بين 1675 م و1725 م؛ وكان أحدهم من أصل من غرب إفريقيا ويحمل المجموعات الفردانية E1b1a-CTS2447 و L3e3b، وكان آخر من أصول ناطقة بلغة البانتو في غرب وسط إفريقيا ويحمل E1b1a-Z5974 و L0a1a2، وكان آخر من أصل من غرب إفريقيا وشرق إفريقيا ويحمل E1b1a-Z5974 و L3d2.[23]
في مقبرة الأمريكيين من أصل أفريقي في فرن كاتوكتين، في فرن كاتوكتين بولاية ماريلاند، تم العثور على 27 أمريكيًا من أصل أفريقي يرجع تاريخهم إلى ما بين 1774 م و1850 م.[24] [25]
كان أحد الذكور الذي كان 98.14% من أصول من جنوب الصحراء الأفريقية يحمل المجموعات الفردانية E1b1a1a1a1c2c و L2a1+143+@16309.[26]
كان أحد الذكور الذي كان 83.73% من أصول أفريقية جنوب الصحراء و7.74% من أصول أوروبية يحمل المجموعات الفردانية E1b1a1a1a1c1b1 و L3e2a1b1.[26]
كان أحد الذكور الذي كان 84.94% من أصول أفريقية جنوب الصحراء الكبرى و9.45% من أصول أوروبية يحمل المجموعات الفردانية E1b1a1a1a2a1a و L2a1+143+16189 (16192)+@16309.[26]
كان أحد الذكور الذي كان 87.83% من أصول أفريقية جنوب الصحراء و8.23% من أصول أوروبية يحمل المجموعات الفردانية E1b1a1a1a1c1a1a3a1d1 و L3d1b3.[26]
كان أحد الذكور الذي كان 98.14% من أصل أفريقي جنوب الصحراء الكبرى يحمل المجموعات الفردانية E1b1a1a1a1a و L3e2a1b1.[26]
كان أحد الذكور الذي كان 93.87% من أصول أفريقية جنوب الصحراء الكبرى و2.58% من أصول أوروبية يحمل المجموعتين الفردانيتين E1b1a1a1 و L3e1.[26]
كان أحد الذكور الذي كان 98.70% من أصول من أفريقيا جنوب الصحراء الكبرى يحمل المجموعات الفردانية E1b1a1a1a1c1b2a و L2a1a1.[26]
كان أحد الذكور الذي كان 97.01% من أصل أفريقي جنوب الصحراء الكبرى يحمل المجموعات الفردانية E1b1a1a1a1c1a1 و L3e2a1b1.[26]
كان أحد الذكور الذي كان 82.31% من أصول أفريقية جنوب الصحراء الكبرى و10.24% من أصول أوروبية يحمل المجموعتين الفردانيتين E1b1a1a1a1c1b و L3e2a1b1.[26]
كان أحد الذكور الذي كان 91.82% من أصول أفريقية جنوب الصحراء الكبرى و5.31% من أصول أوروبية يحمل المجموعات الفردانية E1b1a1a1a1c1a1 و L3e2.[26]
كان أحد الذكور الذي كان 81.18% من أصول أفريقية جنوب الصحراء الكبرى و14.86% من أصول أوروبية يحمل المجموعتين الفردانيتين E1b1a1~ و L2c.[26]
في موقع دفن شارع أنسون في تشارلستون، كارولينا الجنوبية، تم العثور على 18 أمريكيًا من أصل إفريقي يعود تاريخهم إلى القرن الثامن عشر الميلادي.[27]
كان بانزا من أصول غرب وسط إفريقيا يحمل المجموعات الفردانية E1b1a-CTS668 و L3e3b1.[27]
كانت ليما من أصل غرب إفريقي تحمل المجموعتين الفردانيتين E1b1a-M4671 و L3b3.[27]
كان كوتو من أصول غرب وسط إفريقيا يحمل المجموعتين الفردانيتين E1b1a-CTS2198 و L2a1a2.[27]
كانت أنيكا من أصل إفريقي جنوب الصحراء الكبرى تحمل المجموعتين الفردانيتين E1b1a-CTS6126 و L2b1.[27]
كانت نانا من أصل غرب إفريقي تحمل المجموعة الفردانية L2b3a.[27]
كان زيمبو من أصول غرب وسط إفريقيا يحمل المجموعات الفردانية E1b1a-CTS5497 و L3e1e.[27]
كان ووتا من أصول أفريقية جنوب الصحراء الكبرى يحمل المجموعتين الفردانيتين E1b1a-CTS7305 و L3e2b+152.[27]
كان دابا من أصول غرب أفريقي يحمل المجموعات الفردانية E1b1a-M4273 و L2c.[27]
كان فومو من أصول أفريقية جنوب الصحراء الكبرى يحمل المجموعات الفردانية B2a1a-Y12201 و L3e2b+152.[27]
كانت ليزا من أصل غرب أفريقي تحمل المجموعتين الفردانيتين E1b1a-Z6020 و H100.[27]
كان غاندا من أصول غرب أفريقي يحمل المجموعتين الفردانيتين E1b1a-CTS5612 و L1c1c.[27]
كان كوساو من أصول غرب أفريقية وأمريكية أصلية يحمل المجموعتين الفردانيتين E2b1a-CTS2400 و A2.[27]
كان كيدزيرا من أصول غربي وسط أفريقيا يحمل المجموعة الفردانية L2a1a2c.[27]
كان بيتا من أصول أفريقي جنوب الصحراء الكبرى يحمل المجموعات الفردانية E1b1a-M4287 و L3e2b.[27]
كان تيما من أصول غربية وسطية إفريقية يحمل المجموعة الفردانية L3e1e.[27]
كان جود من أصول أفريقية جنوب الصحراء الكبرى يحمل المجموعتين الفردانيتين E1b1a-CTS4975 و L2a1a2c.[27]
كانت أجانا من أصل غربي وسط أفريقيا تحمل المجموعة الفردانية L2a1I.[27]
كان إيسي من أصول غرب وسط أفريقيا يحمل المجموعة الفردانية L3e2a.[27]
الحمض النووي الطبي
تأثير داء الخلايا المنجلية
وسط الصحراء الخضراء، نشأت طفرة الخلايا المنجلية في الصحراء الكبرى[28] أو في منطقة الغابات الشمالية الغربية في غرب إفريقيا الوسطى (على سبيل المثال، الكاميرون)[28] [29] قبل 7300 سنة على الأقل، [28] [29] ] [29] على الرغم من أنه ربما يعود تاريخه إلى 22000 عام مضت.[30] [29]
ربما يكون النمط الفرداني للخلية المنجلية السلفية إلى الأنماط الفردانية الحديثة (على سبيل المثال، أنماط الكاميرون / جمهورية أفريقيا الوسطى وبنين / السنغال ) قد نشأ لأول مرة في أسلاف سكان غرب إفريقيا المعاصرين، الذين يحملون المجموعات الفردانية E1b1a1-L485 و E1b1a1-U175 أو المجموعة الفردانية لأسلافهم E1b1a1-M4732.[28]
ربما هاجر سكان غرب إفريقيا (مثل اليوروبا وإيسان النيجيريين)، الذين يحملون النمط الفرداني للخلية المنجلية في بنين، عبر المنطقة الشمالية الشرقية من إفريقيا إلى المنطقة الغربية من شبه الجزيرة العربية.[28]
من الممكن أن يكون سكان غرب إفريقيا (مثل ميندي في سيراليون)، الحاملين للنمط الفرداني السنغالي للخلية المنجلية، [31] [28] قد هاجروا إلى موريتانيا (77% معدل حدوث حديث) والسنغال (100%)؛ وربما هاجروا أيضًا عبر الصحراء الكبرى، إلى شمال إفريقيا، ومن شمال إفريقيا إلى جنوب أوروبا وتركيا ومنطقة قريبة من شمال العراق وجنوب تركيا.[31]
ربما هاجر البعض وأدخلوا الأنماط الفردية للخلايا المنجلية في السنغال وبنين إلى البصرة في العراق، حيث يحدث كلاهما بالتساوي.[31]
قد يكون سكان غرب إفريقيا، الذين يحملون النمط الفرداني للخلية المنجلية من جنس الذكور، قد هاجروا إلى المنطقة الشمالية من العراق (69.5٪)، والأردن (80٪)، ولبنان (73٪)، وعمان (52.1٪)، ومصر (80.8٪).[31]
التوزيع
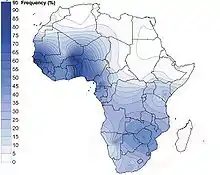
تردد وتنوع E-M2 هو الأعلى في غرب إفريقيا.
أما في داخل إفريقيا، يعرض E-M2 توزيعًا سريريًا من الغرب إلى الشرق ومن الجنوب إلى الشمال.
بمعنى آخر، يتناقص تواتر المجموعة الفردانية مع تحرك الشخص من غرب وجنوب إفريقيا نحو الأجزاء الشرقية والشمالية من إفريقيا.[32]
| المجموعة السكانية | تكرار | مراجع |
|---|---|---|
| باميليكي | 96%-100% | [32] [33] |
| نعجة | 97% | [34] |
| جا | 97% | [34] |
| الهوتو | 94.2% | [32] |
| اليوروبا | 93.1% | [35] |
| التوتسي | 32%-48% | [32] |
| فانتي | 84% | [34] |
| ماندينكا | 79%-87% | [36] [34] |
| أوفامبو | 82% | [34] |
| سنغالي | 81% | [37] |
| غاندا | 77% | [34] |
| بيجاغوس | 76% | [36] |
| بالانتا | 73% | [36] |
| فولا | 73% | [36] |
| كيكويو | 73% | [34] |
| هيريرو | 71% | [34] |
| نالو | 71% | [36] |
تم اختبار السكان في شمال غرب إفريقيا ووسط شرق إفريقيا ومدغشقر وظهرت النتائج بترددات أكثر اعتدالًا.
| المجموعة السكانية | تكرار | مراجع |
|---|---|---|
| الطوارق من تانوت، النيجر | 44.4% (8/18 مادة) | [38] |
| الشيرازي القمري | 41% | [39] |
| الطوارق من جوروم جوروم، بوركينا فاسو | 16.6% (18/3) | [38] |
| الطوارق من جوسي، مالي | 9.1% (1/9) | [38] |
| الرأس الأخضر | 15.9% (201/32) | [40] |
| الماساي | 15.4% (4/26) | [34] |
| لو | 66% (6/9) | [34] |
| العراقو | 11.11% (1/9) | [34] |
| جزر القمر | 23.46% (69/294) | [39] |
| شعب ميرينا (يُطلق عليهم أيضًا سكان المرتفعات) | 44% (4/9) | [41] |
| أنتاندروي | 69.6% (32/46) | [41] |
| أنتانوسي | 48.9% (23/47) | [41] |
| أنتايساكا | 37.5% (3/8) | [41] |
تم العثور على E-M2 على ترددات منخفضة إلى متوسطة في شمال إفريقيا وشمال شرق إفريقيا.
من المحتمل أن تكون بعض الأنساب الموجودة في هذه المناطق بسبب توسع البانتو أو الهجرات الأخرى.[32] [42]
ومع ذلك، فإن اكتشاف علامة E-M2 في عام 2011 م والتي تسبق E-M2 قاد ترومبيتا وآخرين للإشارة إلى أن E-M2 ربما نشأ في شرق إفريقيا.[43]
في إريتريا ومعظم إثيوبيا (باستثناء الأنواك)، عادةً ما يتم العثور على E-V38 في شكل E-M329، وهو أصلي، بينما يشير E-M2 عمومًا إلى أصول البانتو المهاجرة.[44] [45] [46]
| المجموعة السكانية | تكرار | مراجع |
|---|---|---|
| الطوارق من العوينات والتهالة بليبيا | 46.5% (20/43) [arabic-abajed 1] | [47] |
| وهران، الجزائر | 8.6% (8/93) | [48] |
| البربر، جنوب وشمال وسط المغرب | 9.5% (6/6) 5.8% (4/69) | [49] [arabic-abajed 2] [50] |
| العرب المغاربة | 6.8% (3/44) 1.9% (1/54) | [49] [50] |
| صحراويون | 3.5% (29/1) | [49] |
| المصريون | 1.4% (2/147)، 0% (0/73)، 8.33% (3/36) | [32] [51] [52] |
| التونسيون | 1.4% (2/148) | [52] |
| السودانيون (قد يشملون مهاجري الهوسا) | 0.9% (4/445) | [53] |
| المواطنون الصوماليون (قد يشملون أقليات البانتو) | 1.5% (3/201) | [42] |
خارج إفريقيا، تم العثور على E-M2 بترددات منخفضة.
تم العثور على E-M2 بترددات منخفضة في غرب آسيا.
كما لوحظت بعض الأحداث المعزولة لـ E-M2 بين السكان في جنوب أوروبا، مثل كرواتيا ومالطا وإسبانيا والبرتغال.[54] [55] [56] [57]
| المجموعة السكانية | تكرار | مراجع |
|---|---|---|
| بحرينيين | 8.6% (46/562) | [58] |
| سعوديون | 6.6% (11/157) | [59] |
| عمانيون | 6.6% (8/121) | [32] |
| إماراتيون | 5.5% (9/164) | [60] |
| يمنيون | 4.8% (3/62) | [60] |
| القبارصة | 3.2% (2/62) | [57] |
| الإيرانيين الجنوبيين | 1.7% (2/117) | [61] |
| أردنيون | 1.4% (2/139) | [62] |
| سيريلانكا | 1.4% (9/638) | [63] |
| الجزر الإيولية، إيطاليا | 1.2% (1/81) | [64] |
جلبت تجارة العبيد عبر المحيط الأطلسي الشعب إلى أمريكا الشمالية وأمريكا الوسطى وأمريكا الجنوبية بما في ذلك منطقة البحر الكاريبي.
وبالتالي، غالبًا ما تتم ملاحظة المجموعة الفردانية في سكان الولايات المتحدة عند الرجال الذين يعرفون أنفسهم على أنهم أمريكيون من أصل أفريقي.[65]
وقد لوحظ أيضًا في عدد من السكان في المكسيك ومنطقة البحر الكاريبي وأمريكا الوسطى وأمريكا الجنوبية بين الأشخاص من أصل أفريقي.
| المجموعة السكانية | تكرار | مراجع |
|---|---|---|
| الأمريكيون | 7.7–7.9% [arabic-abajed 3] | [65] |
| الكوبيين | 9.8% (13/132) | [66] |
| الدومينيكان | 5.69% (2/26) | [67] |
| البورتوريكيين | 19.23% (5/26) | [67] |
| نيكاراغوا | 5.5% (9/165) | [68] |
| عدة مجموعات من الكولومبيين | 6.18% (69/1116) | [69] |
| ألاغواس، البرازيل | 4.45% (11/247) | [70] |
| باهيا، البرازيل | 19% (19/100) | [71] |
| جزر البهاما | 58.63% (251/428) | [72] |
الفئات الفرعية
E1b1a1
يتم تعريف E1b1a1 بالعلامات التالية:
DYS271/M2/SY81 وM291 وP1/PN1 وP189 وP293 وV43 وV95.
بينما يصل E1b1a إلى أعلى تردد له وهو 81% في السنغال، أظهر 1 فقط من أصل 139 سنغاليًا تم اختبارهم M191/P86،[37] وبعبارة أخرى، عندما ينتقل المرء إلى غرب إفريقيا من غرب إفريقيا الوسطى، يتم العثور على الفئة الفرعية الأقل E1b1a1f.
كروسياني (2002) يقول: "قد يكون التفسير المحتمل هو أن كروموسومات النمط الفرداني 24 [E-M2*] كانت موجودة بالفعل عبر الحزام السوداني عندما ظهرت طفرة M191، التي تحدد النمط الفرداني 22، في وسط غرب أفريقيا، عندها فقط قد يؤدي التوسع الديموغرافي اللاحق إلى جلب كروموسومات النمط الفرداني 22 من وسط غرب أفريقيا إلى غرب أفريقيا، مما يؤدي إلى ظهور توزيعات سريرية معاكسة للنمطين الفردانيين 22 و24."[33]
E1b1a1a1
يتم تعريف E1b1a1a1 بشكل شائع بـ M180/P88.
تتم ملاحظة الطبقة الفرعية القاعدية بانتظام في عينات M2+.
E1b1a1a1a
يتم تعريف E1b1a1a1a بواسطة العلامة M58. 5% (2/37) من مدينة سينجا ريمايبي، بوركينا فاسو كانت نتيجة اختبارها إيجابية لـ E-M58. [33] 15% (10/69) من الهوتو في رواندا ثبتت إصابتهم بـ M58.[32]
وقد ثبتت إصابة ثلاثة مواطنين من جنوب إفريقيا بهذه العلامة.[73] كانت نتيجة اختبار كاريوكا واحدة من ريو دي جانيرو بالبرازيل إيجابية بالنسبة لـ M58 SNP.[74] لم يتم الإبلاغ عن مكان المنشأ والعمر.
E1b1a1a1b
يتم تعريف E1b1a1a1b بواسطة M116.2، وهي علامة خاصة.
وتم العثور على ناقلة واحدة في مالي. [73] [arabic-abajed 4]
E1b1a1a1c
يتم تعريف E1b1a1a1c بواسطة علامة خاصة M149.
تم العثور على هذه العلامة في جنوب إفريقيا. [73]
E1b1a1a1d
يتم تعريف E1b1a1a1d بواسطة علامة خاصة وهي M155.
E1b1a1a1e
يتم تعريف E1b1a1a1e بواسطة علامات M10 وM66 وM156 وM195. كان اختبار شعب ويراك في تنزانيا إيجابيًا بنسبة 4.6% (2/43) لفيروس E-M10.[75]
تم العثور على E-M10 لدى شخص واحد من مجموعة ليسونغو في جمهورية إفريقيا الوسطى وعضوين من مجموعة سكانية "مختلطة" من منطقة أداماوا.[76]
E1b1a1a1f
يتم تعريف E1b1a1a1f بـ L485.
يبدو أن العقدة القاعدية E-L485* غير شائعة إلى حد ما ولكن لم يتم اختبارها بشكل كافٍ على أعداد كبيرة من السكان.
تم اكتشاف L485 SNP السلفي (إلى جانب العديد من فئاته الفرعية) مؤخرًا.
بعض هذه SNPs لديها القليل من البيانات السكانية المنشورة أو لا تحتوي على أي بيانات سكانية منشورة و/أو لم تحصل بعد على اعتراف بالتسميات من قبل YCC.
- يتم تعريف E1b1a1a1f1 بواسطة العلامة L514. لا يحتوي هذا SNP حاليًا على بيانات دراسة سكانية خارج مشروع 1000 جينوم.
- يتم تعريف E1b1a1a1f1a (YCC E1b1a7) بواسطة العلامة M191/P86. فيليبو وآخرون. (2011) درس عددًا من السكان الأفارقة الذين كانوا إيجابيين لـ E-M2 ووجدوا E-M191/P86 الأساسي (بدون E-P252/U174) في مجموعة من المتحدثين باللغة الغورية في بوركينا فاسو. [77] مونتانو وآخرون. (2011) وجد توزيعًا متفرقًا مشابهًا لـ E-M191* في نيجيريا والجابون والكاميرون والكونغو. [78] حدثت عينات إيجابية لفيروس M191/P86 في المجموعات السكانية التي تم اختبارها في أنانج (38.3%)، وإيبيبيو (45.6%)، وإفيك (45%)، وإيغبو (54.3%) الذين يعيشون في نيجيريا، غرب أفريقيا. [79] يظهر E-M191/P86 بترددات مختلفة في وسط وجنوب أفريقيا ولكن جميعها تقريبًا إيجابية أيضًا لـ P252/U174. كان اختبار جنوب إفريقيا الناطقين بالبانتو (89/343) إيجابيًا بنسبة 25.9٪، وكان اختبار جنوب إفريقيا الناطقين بالخوي-سان إيجابيًا بنسبة 7.7٪ (14/183) لهذا SNP. [80] كما يظهر بشكل شائع عند الأفارقة الذين يعيشون في الأمريكتين. كان اختبار السكان في ريو دي جانيرو بالبرازيل إيجابيًا بنسبة 9.2٪ (12/130). [81] 34.9% (29/83) من الرجال الأمريكيين من أصل أفريقي كانت نتيجة اختبارهم إيجابية لمرض M191. [82]
- فيراما وآخرون. (2010) تشير الدراسات التي أجريت على أجزاء إعادة تجميع كروموسومات Y الإيجابية M191 إلى أن هذا النسب "انتشر بشكل منتشر مع أنماط فردية متعددة عالية التردد مما يشير إلى فترة تطورية أطول منذ ظهور هذه المجموعة الفردانية". [83] يبدو أن الفئة الفرعية E1b1a1a1f1a تعبر عن التوزيعات السريرية المعاكسة لـ E1b1a1 * في منطقة السافانا بغرب إفريقيا. المجموعة الفردانية E1b1a1a1f1a (E-M191) لها تردد 23% في الكاميرون (حيث تمثل 42% من الأنماط الفردانية التي تحمل طفرة DYS271 أو E-M2)، و13% في بوركينا فاسو (16% من الأنماط الفردية التي تحمل طفرة M2/DYS271). و1% فقط في السنغال.[84] وبالمثل، في حين يصل E1b1a إلى أعلى تردد له وهو 81٪ في السنغال، فإن 1 فقط من أصل 139 سنغاليًا تم اختبارهم أظهروا M191/P86. [37] وبعبارة أخرى، عندما ينتقل المرء إلى غرب أفريقيا من غرب أفريقيا الوسطى، يتم العثور على الفئة الفرعية الأقل E1b1a1f. "قد يكون التفسير المحتمل هو أن كروموسومات النمط الفرداني 24 [E-M2*] كانت موجودة بالفعل عبر الحزام السوداني عندما ظهرت طفرة M191، التي تحدد النمط الفرداني 22، في وسط غرب إفريقيا. عندها فقط كان التوسع الديموغرافي اللاحق قد جلب كروموسومات النمط الفرداني 22 من وسط غرب أفريقيا إلى غرب أفريقيا، مما أدى إلى ظهور توزيعات سريرية معاكسة للنمطين الفردانيين 22 و24." [85]
- يتم تعريف E1b1a1a1f1a1 (YCC E1b1a7a) بواسطة P252/U174. يبدو أنها الفئة الفرعية الأكثر شيوعًا من E-L485. ويعتقد أنه نشأ بالقرب من غرب أفريقيا الوسطى. ونادرا ما توجد في الأجزاء الغربية من غرب أفريقيا. مونتانو وآخرون. (2011) وجد أن هذه الفئة الفرعية منتشرة جدًا في نيجيريا والجابون.[86] فيليبو وآخرون. (2011) قدر tMRCA بحوالي 4.2 كيا من عينة من سكان اليوروبا إيجابيين بالنسبة للـ SNP.[87]
- يتم تعريف E1b1a1a1f1a1b (YCC E1b1a7a2) بواسطة P115. تمت ملاحظة هذه الفئة الفرعية فقط بين شعب فانغ في وسط أفريقيا.[78]
- يتم تعريف E1b1a1a1f1a1c (YCC E1b1a7a3) بواسطة P116. مونتانو وآخرون. (2011) لاحظ هذا النمط SNP فقط في الجابون وسكان الباسا من الكاميرون.[78]
- يتم تعريف E1b1a1a1f1a1d بواسطة Z1704. وقد لوحظت هذه الفئة الفرعية في جميع أنحاء أفريقيا. وجد اتحاد مشروع 1000 جينوم هذا SNP في يوروبا نيجيري، وثلاثة لوهايا كينية وواحد من أصل أفريقي بورتوريكو.[88]
- يتم تعريف E1b1a1a1f1b بواسطة العلامات L515 وL516 وL517 وM263.2. تم العثور على هذه الفئة الفرعية من قبل الباحثين في مشروع مقارنة جينوم Y-Chromosome باستخدام بيانات من شركة المعلوماتية الحيوية التجارية 23andMe.[89]
E1b1a1a1g
يتم تعريف E1b1a1a1g (YCC E1b1a8) بواسطة العلامة U175.
يعتبر E-U175* الأساسي نادرًا للغاية.
مونتانو وآخرون. (2011) وجد واحدًا فقط من بين 505 من الأشخاص الأفارقة الذين تم اختبارهم والذي كان إيجابيًا لـ U175 ولكنه سلبي لـ U209.[86]
بروكاتو وآخرون. وجدت ترددات منخفضة مماثلة من القاعدية E-U175 * في مواضيع في ساحل العاج وبنين. فيراما وآخرون. (2010) وجد U175 في أنانج المختبر (45.3%)، إيبيبيو (37%)، إيفيك (33.3%)، وإجبو (25.3%) لكنه لم يختبر U209.[83]
إن "النمط الفرداني البانتو" المفترض الموجود في حاملات E-U175 "موجود بترددات ملحوظة في لغات النيجر والكونغو الأخرى التي تتحدثها الشعوب في أقصى الغرب مثل غينيا بيساو ".[83] هذا هو النمط الفرداني المشروط لعلامات STR الشائعة في حاملات E-U175.[arabic-abajed 5]
| النمط الفرداني E-U175 | DYS19 | DYS388 | DYS390 | DYS391 | DYS392 | DYS393 | |
| 15 | 12 | 21 | 10 | 11 | 13 | ||
يحتوي E1b1a1a1g على عدة فئات فرعية.
- يتم تعريف E1b1a1a1g1 (YCC E1b1a8a) بواسطة U209. إنها الفئة الفرعية الأبرز في U175. هذه الفئة الفرعية لها ترددات عالية جدًا تزيد عن خمسين بالمائة في السكان الكاميرونيين في باسا وباكاكا، مما قد يشير إلى مكان المنشأ. ومع ذلك، يتم العثور على E-U209 على نطاق واسع في الترددات المنخفضة في بلدان غرب ووسط أفريقيا المحيطة بالكاميرون والجابون.[86] بروكاتو وآخرون. (2010) وجد الحزب الوطني الاسكتلندي في سكان أهيزي (في ساحل العاج ) 38.8% (19/49)، ويعقوبا (ساحل العاج) 27.5% (11/40)، والبنينيين 6.5% (5/77) على التوالي.[90]
- يتم تعريف E1b1a1a1g1a (YCC E1b1a8a1) بواسطة U290. مونتانو وآخرون. (2011) أظهرت دراسة U290 ترددًا أقل في نيجيريا (11.7٪) وغرب أفريقيا الوسطى مقارنة بالعقدة القاعدية U209. وكان أعلى معدل تكرار للسكان في تلك الدراسة هو 57.7% (15/26) في إواندو في الكاميرون.[78] تم اختبار 32.5% (27/83) من الرجال الأمريكيين من أصل أفريقي بواسطة سيمز وآخرون. (2007) كانت إيجابية لهذا SNP.[82]
- يتم تعريف E1b1a1a1g1a2 بواسطة Z1725. تمت ملاحظة هذه العلامة من قبل اتحاد مشروع الجينوم الـ 1000 في اليوروبا النيجيريين ولوهيا الكينيين.[91]
- يتم تعريف E1b1a1a1g1c (YCC E1b1a4) بواسطة M154. تم اختبار عدد سكان باميليكي بنسبة 31.3٪ (15/48) للعلامة. تم اختبار مكبرات الصوت باكاكا من الكاميرون بنسبة 8%.[85] وجد مجتمع اختبار Ovimbundu هذا SNP بنسبة 14٪ (14/100).[92] تم العثور أيضًا على أعضاء من هذه الفئة الفرعية في جنوب إفريقيا.[93] [94]
- يتم تعريف E1b1a1a1g1d بواسطة V39. ترومبيتا وآخرون. نشرت SNP لأول مرة في عام 2011 ولكنها قدمت القليل من البيانات السكانية عنها.[95] ومن المعروف أنه تم العثور عليه فقط في السكان الأفارقة.
علم الوراثة
التاريخ التطوري
قبل عام 2002 م، كان هناك في الأدبيات الأكاديمية ما لا يقل عن سبعة أنظمة تسمية لشجرة النشوء والتطور للكروموسوم Y. وقد أدى هذا إلى ارتباك كبير.
في عام 2002 م، اجتمعت المجموعات البحثية الرئيسية معًا وشكلت اتحاد كروموسوم Y (YCC).
لقد نشروا ورقة مشتركة أنشأت شجرة واحدة جديدة اتفق الجميع على استخدامها.
وفي وقت لاحق، شكلت مجموعة من العلماء المواطنين المهتمين بعلم الوراثة السكانية وعلم الأنساب الجيني مجموعة عمل لإنشاء شجرة هواة تهدف قبل كل شيء إلى أن تكون في الوقت المناسب.
يجمع الجدول أدناه كل هذه الأعمال عند نقطة شجرة YCC التاريخية لعام 2002.
وهذا يسمح للباحث بمراجعة الأدبيات المنشورة القديمة للتنقل بسرعة بين التسميات.
أنظر أيضًا
علم الوراثة
 بوابة علم الوراثة
بوابة علم الوراثة
- Wang، Ke؛ وآخرون (2020). "Ancient genomes reveal complex patterns of population movement, interaction, and replacement in sub-Saharan Africa". Science Advances. ج. 6 ع. 24: eaaz0183. Bibcode:2020SciA....6..183W. DOI:10.1126/sciadv.aaz0183. ISSN:2375-2548. OCLC:8616876709. PMC:7292641. PMID:32582847.
- Wang، Ke؛ وآخرون (2020). "Supplementary Materials for Ancient genomes reveal complex patterns of population movement, interaction, and replacement in sub-Saharan Africa" (PDF). Science Advances. ج. 6 ع. 24: eaaz0183. Bibcode:2020SciA....6..183W. DOI:10.1126/sciadv.aaz0183. ISSN:2375-2548. OCLC:8616876709. PMC:7292641. PMID:32582847. مؤرشف من الأصل (PDF) في 2023-03-14.
- Hawass، Zahi؛ وآخرون (2012). "Revisiting the harem conspiracy and death of Ramesses III: anthropological, forensic, radiological, and genetic study". British Medical Journal. ج. 345: e8268. DOI:10.1136/bmj.e8268. ISSN:0959-8138. OCLC:825973553. PMID:23247979. مؤرشف من الأصل في 2023-03-07.
- Gad، Yehia Z؛ وآخرون (2021). "Insights from ancient DNA analysis of Egyptian human mummies: clues to disease and kinship". Human Molecular Genetics. ج. 30 ع. R1: R24–R28. DOI:10.1093/hmg/ddaa223. ISSN:0964-6906. OCLC:8681412353. PMID:33059357.
- Prendergast، Mary E.؛ وآخرون (30 مايو 2019). "Ancient DNA reveals a multistep spread of the first herders into sub-Saharan Africa". Science. ج. 365 ع. 6448. Bibcode:2019Sci...365.6275P. DOI:10.1126/science.aaw6275. ISSN:0036-8075. OCLC:8176642048. PMC:6827346. PMID:31147405.
- Prendergast، Mary E.؛ وآخرون (30 مايو 2019). "Supplementary Materials for Ancient DNA reveals a multistep spread of the first herders into sub-Saharan Africa" (PDF). Science. ج. 365 ع. 6448. Bibcode:2019Sci...365.6275P. DOI:10.1126/science.aaw6275. ISSN:0036-8075. OCLC:8176642048. PMC:6827346. PMID:31147405. مؤرشف من الأصل (PDF) في 2022-10-08.
- Brielle، Esther S.؛ وآخرون (29 مارس 2023). "Supplementary Data Files for Entwined African and Asian genetic roots of medieval peoples of the Swahili coast". Nature. ج. 615 ع. 7954: 866–873. Bibcode:2023Natur.615..866B. DOI:10.1038/s41586-023-05754-w. ISSN:0028-0836. OCLC:9819552636. PMC:10060156. PMID:36991187.
- Barquera، Rodrigo؛ وآخرون (8 يونيو 2020). "Origin and Health Status of First-Generation Africans from Early Colonial Mexico". Current Biology. ج. 30 ع. 11: 2078–2091. DOI:10.1016/j.cub.2020.04.002. ISSN:0960-9822. OCLC:8586564917. PMID:32359431.
- Fortes-Lima، Cesar A. (22 نوفمبر 2021). "Disentangling the Impact of the Transatlantic Slave Trade in African Diaspora Populations from a Genomic Perspective". Africa, the Cradle of Human Diversity Cultural and Biological Approaches to Uncover African Diversity. Brill. ص. 305, 308–321. DOI:10.1163/9789004500228_012. ISBN:978-90-04-50022-8. OCLC:1284920909.
- Peyroteo-Stjerna، Rita؛ وآخرون (21 فبراير 2022). "Multidisciplinary investigation reveals an individual of West African origin buried in a Portuguese Mesolithic shell midden four centuries ago". Journal of Archaeological Science: Reports. ج. 42: 103370. DOI:10.1016/j.jasrep.2022.103370. OCLC:1337974923.
- Sandoval-Velasco، Marcela؛ وآخرون (7 سبتمبر 2023). "The ancestry and geographical origins of St Helena's liberated Africans" (PDF). American Journal of Human Genetics. ج. 110 ع. 9: 1590–1599. DOI:10.1016/j.ajhg.2023.08.001. ISSN:0002-9297. OCLC:9998699240. PMID:37683613. مؤرشف من الأصل (PDF) في 2023-09-14.
- Isable، Kendra Briana (أغسطس 2021). "Contributing to the Discussion: The Health of Enslaved Africans Through the Lens of Bioarchaeology". California State University, Northridge. ص. 34. مؤرشف من الأصل في 2023-09-14.
- Fortes-Lima، Cesar؛ Verdu، Paul (17 ديسمبر 2020). "Anthropological genetics perspectives on the transatlantic slave trade". Human Molecular Genetics. ج. 30 ع. R1: R79–R87. DOI:10.1093/hmg/ddaa271. PMID:33331897. مؤرشف من الأصل في 2023-03-12.
- Callaway، Ewen (7 ديسمبر 2016). "What DNA reveals about St Helena's freed slaves". Nature. ج. 540 ع. 7632: 184–187. DOI:10.1038/540184a.
- Sandoval-Velasco، Marcela؛ وآخرون (7 سبتمبر 2023). "Data S1. Tables S1–S13: The ancestry and geographical origins of St Helena's liberated Africans". American Journal of Human Genetics. ج. 110 ع. 9: 1590–1599. DOI:10.1016/j.ajhg.2023.08.001. ISSN:0002-9297. OCLC:9998699240. PMID:37683613. مؤرشف من الأصل في 2023-09-14.
- Fleskes، Raquel E.؛ وآخرون (2019). "Ancient DNA and bioarchaeological perspectives on European and African diversity and relationships on the colonial Delaware frontier". American Journal of Physical Anthropology. ج. 170 ع. 2: 232–245. DOI:10.1002/ajpa.23887. PMID:31270812. مؤرشف من الأصل في 2022-03-07.
- Racimo، Fernando؛ وآخرون (2020). "Beyond broad strokes: sociocultural insights from the study of ancient genomes" (PDF). Nature Reviews Genetics. ج. 21 ع. 6: 355–366. DOI:10.1038/s41576-020-0218-z. ISSN:1471-0056. PMID:32127690. مؤرشف من الأصل (PDF) في 2022-04-14.
- Callaway، Ewen (5 نوفمبر 2019). "Genomes trace origins of enslaved people who died on remote island". Nature. ج. 575 ع. 18: 18. Bibcode:2019Natur.575...18C. DOI:10.1038/d41586-019-03152-9. PMID:31690869. مؤرشف من الأصل في 2023-09-14.
- Abel، Sarah؛ Schroeder، Hannes (أكتوبر 2020). "From Country Marks to DNA Markers: The Genomic Turn in the Reconstruction of African Identities". Current Anthropology. ج. 61: S206. DOI:10.1086/709550. مؤرشف من الأصل في 2022-04-22.
- Olalde، Iñigo؛ وآخرون (2019). "Supplementary Materials for The genomic history of the Iberian Peninsula over the past 8000 years" (PDF). Science. ج. 363 ع. 6432: 1230–1234. Bibcode:2019Sci...363.1230O. DOI:10.1126/science.aav4040. ISSN:0036-8075. OCLC:8024095449. PMC:6436108. PMID:30872528. مؤرشف من الأصل (PDF) في 2022-12-25.
- Olalde، Iñigo (2019). "The genomic history of the Iberian Peninsula over the past 8000 years, TablesS1-S5". Science. ج. 363 ع. 6432: 1230–1234. Bibcode:2019Sci...363.1230O. DOI:10.1126/science.aav4040. ISSN:0036-8075. OCLC:8024095449. PMC:6436108. PMID:30872528.
- Olalde، Iñigo (2019). "Materials/Methods, Supplementary Text, Tables, Figures, and/or References". Science. ج. 363 ع. 6432: 1230–1234. Bibcode:2019Sci...363.1230O. DOI:10.1126/science.aav4040. ISSN:0036-8075. OCLC:8024095449. PMC:6436108. PMID:30872528.
- Fleskes، Raquel E.؛ وآخرون (5 يونيو 2023). "Historical genomes elucidate European settlement and the African diaspora in Delaware" (PDF). Current Biology. ج. 33 ع. 11: 2350-2358.e7. DOI:10.1016/j.cub.2023.04.069. ISSN:0960-9822. OCLC:9874997102. PMID:37207647. مؤرشف من الأصل (PDF) في 2023-06-29.
- Harney، Éadaoin؛ وآخرون (2023). "The genetic legacy of African Americans from Catoctin Furnace" (PDF). ج. 381 ع. 500: eade4995. DOI:10.1126/science.ade4995. PMID:37535739. مؤرشف من الأصل (PDF) في 2023-08-06.
{{استشهاد بدورية محكمة}}: الاستشهاد بدورية محكمة يطلب|دورية محكمة=(مساعدة) - Harney، Éadaoin؛ وآخرون (2023). "Supplementary Materials for The genetic legacy of African Americans from Catoctin Furnace" (PDF). ج. 381 ع. 500: 1–115. DOI:10.1126/science.ade4995. PMID:37535739. مؤرشف من الأصل (PDF) في 2023-08-06.
{{استشهاد بدورية محكمة}}: الاستشهاد بدورية محكمة يطلب|دورية محكمة=(مساعدة) - Harney، Éadaoin؛ وآخرون (2023). "Tables S1 to S24, S1.1, S3.2 to S3.7, and S4.1 for The genetic legacy of African Americans from Catoctin Furnace". ج. 381 ع. 500: eade4995. DOI:10.1126/science.ade4995. PMID:37535739. مؤرشف من الأصل في 2023-08-06.
{{استشهاد بدورية محكمة}}: الاستشهاد بدورية محكمة يطلب|دورية محكمة=(مساعدة) - Fleskes، Raquel E.؛ وآخرون (2023). "Community-engaged ancient DNA project reveals diverse origins of 18th-century African descendants in Charleston, South Carolina". Anthropology. ج. 120 ع. 3: e2201620120. DOI:10.1073/pnas.2201620120. PMC:9934026. PMID:36623185. مؤرشف من الأصل في 2023-02-02.
- Shriner، Daniel؛ Rotimi، Charles N. (2018). "Whole-Genome-Sequence-Based Haplotypes Reveal Single Origin of the Sickle Allele during the Holocene Wet Phase". American Journal of Human Genetics. ج. 102 ع. 4: 547–556. DOI:10.1016/j.ajhg.2018.02.003. ISSN:0002-9297. OCLC:7352712531. PMC:5985360. PMID:29526279.
- Esoh، Kevin؛ Wonkam، Ambroise (2021). "Evolutionary history of sickle-cell mutation: implications for global genetic medicine". Human Molecular Genetics. ج. 30 ع. R1: R119–R128. DOI:10.1093/hmg/ddab004. ISSN:0964-6906. OCLC:8885008275. PMC:8117455. PMID:33461216.
- Laval، Guillaume؛ وآخرون (2019). "Recent Adaptive Acquisition by African Rainforest Hunter-Gatherers of the Late Pleistocene Sickle-Cell Mutation Suggests Past Differences in Malaria Exposure". The American Journal of Human Genetics. ج. 104 ع. 3: 553–561. DOI:10.1016/j.ajhg.2019.02.007. ISSN:0002-9297. OCLC:8015758034. PMC:6407493. PMID:30827499.
- Yaseen، Noor Taha؛ وآخرون (2020). "Sickle ß-globin haplotypes among patients with sickle cell anemia in Basra, Iraq: A cross-sectional study". Iraqi Journal of Hematology. ج. 9 ع. 1: 23–29. DOI:10.4103/ijh.ijh_20_19. ISSN:2072-8069. OCLC:8663256900. مؤرشف من الأصل في 2023-06-03.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - "The Levant versus the Horn of Africa: evidence for bidirectional corridors of human migrations". American Journal of Human Genetics. ج. 74 ع. 3: 532–44. مارس 2004. DOI:10.1086/382286. PMC:1182266. PMID:14973781.
- "A back migration from Asia to sub-Saharan Africa is supported by high-resolution analysis of human Y-chromosome haplotypes". American Journal of Human Genetics. ج. 70 ع. 5: 1197–214. مايو 2002. DOI:10.1086/340257. PMC:447595. PMID:11910562.
- "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes". European Journal of Human Genetics. ج. 13 ع. 7: 867–76. يوليو 2005. DOI:10.1038/sj.ejhg.5201408. PMID:15856073.
- International HapMap Consortium (أكتوبر 2005). "A haplotype map of the human genome". Nature. ج. 437 ع. 7063: 1299–320. Bibcode:2005Natur.437.1299T. DOI:10.1038/nature04226. PMC:1880871. PMID:16255080.
- "Y-chromosomal diversity in the population of Guinea-Bissau: a multiethnic perspective". BMC Evolutionary Biology. ج. 7: 124. يوليو 2007. DOI:10.1186/1471-2148-7-124. PMC:1976131. PMID:17662131.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - "Ethiopians and Khoisan share the deepest clades of the human Y-chromosome phylogeny". American Journal of Human Genetics. ج. 70 ع. 1: 265–8. يناير 2002. DOI:10.1086/338306. PMC:384897. PMID:11719903.
- "Linking the sub-Saharan and West Eurasian gene pools: maternal and paternal heritage of the Tuareg nomads from the African Sahel". European Journal of Human Genetics. ج. 18 ع. 8: 915–23. أغسطس 2010. DOI:10.1038/ejhg.2010.21. PMC:2987384. PMID:20234393.
- "Genetic diversity on the Comoros Islands shows early seafaring as major determinant of human biocultural evolution in the Western Indian Ocean". European Journal of Human Genetics. ج. 19 ع. 1: 89–94. يناير 2011. DOI:10.1038/ejhg.2010.128. PMC:3039498. PMID:20700146.
- "Y-chromosome lineages in Cabo Verde Islands witness the diverse geographic origin of its first male settlers". Human Genetics. ج. 113 ع. 6: 467–72. نوفمبر 2003. DOI:10.1007/s00439-003-1007-4. PMID:12942365.
- "On the origins and admixture of Malagasy: new evidence from high-resolution analyses of paternal and maternal lineages". Molecular Biology and Evolution. ج. 26 ع. 9: 2109–24. سبتمبر 2009. DOI:10.1093/molbev/msp120. PMID:19535740.
- "High frequencies of Y chromosome lineages characterized by E3b1, DYS19-11, DYS392-12 in Somali males". European Journal of Human Genetics. ج. 13 ع. 7: 856–66. يوليو 2005. DOI:10.1038/sj.ejhg.5201390. PMID:15756297.
- MacAulay، Vincent، المحرر (يناير 2011). "A new topology of the human Y chromosome haplogroup E1b1 (E-P2) revealed through the use of newly characterized binary polymorphisms". PLOS ONE. ج. 6 ع. 1: e16073. Bibcode:2011PLoSO...616073T. DOI:10.1371/journal.pone.0016073. PMC:3017091. PMID:21253605.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - "Forensic data and microvariant sequence characterization of 27 Y-STR loci analyzed in four Eastern African countries". Forensic Science International. Genetics. ج. 27: 123–131. مارس 2017. DOI:10.1016/j.fsigen.2016.12.015. PMID:28068531.
- Plaster et al. Y-DNA E subclades نسخة محفوظة 2023-09-04 على موقع واي باك مشين.
- Variation in Y chromosome, mitochondrial DNA and labels of identity on Ethiopia (Doctoral thesis). 28 سبتمبر 2011. مؤرشف من الأصل في 2023-05-26.
- "Deep into the roots of the Libyan Tuareg: a genetic survey of their paternal heritage". American Journal of Physical Anthropology. ج. 145 ع. 1: 118–24. مايو 2011. DOI:10.1002/ajpa.21473. PMID:21312181.
- "Analysis of Y-chromosomal SNP haplogroups and STR haplotypes in an Algerian population sample". International Journal of Legal Medicine. ج. 122 ع. 3: 251–5. مايو 2008. DOI:10.1007/s00414-007-0203-5. PMID:17909833.
- "High-resolution analysis of human Y-chromosome variation shows a sharp discontinuity and limited gene flow between northwestern Africa and the Iberian Peninsula". American Journal of Human Genetics. ج. 68 ع. 4: 1019–29. أبريل 2001. DOI:10.1086/319521. PMC:1275654. PMID:11254456.
- "Phylogeographic analysis of haplogroup E3b (E-M215) y chromosomes reveals multiple migratory events within and out of Africa". American Journal of Human Genetics. ج. 74 ع. 5: 1014–22. مايو 2004. DOI:10.1086/386294. PMC:1181964. PMID:15042509.
- "Ancestral Asian source(s) of new world Y-chromosome founder haplotypes". American Journal of Human Genetics. ج. 64 ع. 3: 817–31. مارس 1999. DOI:10.1086/302282. PMC:1377800. PMID:10053017.
- "A predominantly neolithic origin for Y-chromosomal DNA variation in North Africa". American Journal of Human Genetics. ج. 75 ع. 2: 338–45. أغسطس 2004. DOI:10.1086/423147. PMC:1216069. PMID:15202071.
- "Y-chromosome variation among Sudanese: restricted gene flow, concordance with language, geography, and history". American Journal of Physical Anthropology. ج. 137 ع. 3: 316–23. نوفمبر 2008. DOI:10.1002/ajpa.20876. PMID:18618658.
- Branka Grskovic, Andro Vrdoljak, Maja Popovic, Ivica Valpotic, Simun Andelinovic, Vlastimil Stenzl, Edvard Ehler, Ludvik Urban, Gordana Lackovic, Peter Underhill, Dragan Primorac. "Croatian national reference Y-STR haplotype database". Molecular Biology Reports. ج. 39 ع. 7: 7727–41. يوليو 2012. DOI:10.1007/s11033-012-1610-3. PMID:22391654.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: آخرون (link) - "Population structure in the Mediterranean basin: a Y chromosome perspective". Annals of Human Genetics. ج. 70 ع. Pt 2: 207–25. مارس 2006. DOI:10.1111/j.1529-8817.2005.00224.x. PMID:16626331.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Reduced genetic structure of the Iberian peninsula revealed by Y-chromosome analysis: implications for population demography". European Journal of Human Genetics. ج. 12 ع. 10: 855–63. أكتوبر 2004. DOI:10.1038/sj.ejhg.5201225. PMID:15280900.
- "The genetic legacy of religious diversity and intolerance: paternal lineages of Christians, Jews, and Muslims in the Iberian Peninsula". American Journal of Human Genetics. ج. 83 ع. 6: 725–36. ديسمبر 2008. DOI:10.1016/j.ajhg.2008.11.007. PMC:2668061. PMID:19061982.
- Noora R. Al-Snan1؛ Safia A. Messaoudi؛ Yahya M. Khubrani؛ Jon H. Wetton؛ Mark A. Jobling؛ Moiz Bakhiet (2020). "Geographical structuring and low diversity of paternal lineages in Bahrain shown by analysis of 27 Y-STRs". Molecular Genetics and Genomics. ج. 295 ع. 6: 1315–1324. DOI:10.1007/s00438-020-01696-4. PMC:7524810. PMID:32588126.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: أسماء عددية: قائمة المؤلفين (link) - "Saudi Arabian Y-Chromosome diversity and its relationship with nearby regions". BMC Genetics. ج. 10: 59. سبتمبر 2009. DOI:10.1186/1471-2156-10-59. PMC:2759955. PMID:19772609.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - "Y-chromosome diversity characterizes the Gulf of Oman". European Journal of Human Genetics. ج. 16 ع. 3: 374–86. مارس 2008. DOI:10.1038/sj.ejhg.5201934. PMID:17928816.
- "Iran: tricontinental nexus for Y-chromosome driven migration". Human Heredity. ج. 61 ع. 3: 132–43. 2006. DOI:10.1159/000093774. PMID:16770078.
- "Y-chromosome and mtDNA polymorphisms in Iraq, a crossroad of the early human dispersal and of post-Neolithic migrations". Molecular Phylogenetics and Evolution. ج. 28 ع. 3: 458–72. سبتمبر 2003. DOI:10.1016/S1055-7903(03)00039-3. PMID:12927131.
- "Y-chromosomal evidence for a limited Greek contribution to the Pathan population of Pakistan". European Journal of Human Genetics. ج. 15 ع. 1: 121–6. يناير 2007. DOI:10.1038/sj.ejhg.5201726. PMC:2588664. PMID:17047675.
- "Excavating Y-chromosome haplotype strata in Anatolia". Human Genetics. ج. 114 ع. 2: 127–48. يناير 2004. DOI:10.1007/s00439-003-1031-4. PMID:14586639.
- "Sub-populations within the major European and African derived haplogroups R1b3 and E3a are differentiated by previously phylogenetically undefined Y-SNPs". Human Mutation. ج. 28 ع. 1: 97. يناير 2007. DOI:10.1002/humu.9469. PMID:17154278.
- "Genetic origin, admixture, and asymmetry in maternal and paternal human lineages in Cuba". BMC Evolutionary Biology. ج. 8: 213. يوليو 2008. DOI:10.1186/1471-2148-8-213. PMC:2492877. PMID:18644108.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - "Colloquium paper: genome-wide patterns of population structure and admixture among Hispanic/Latino populations". Proceedings of the National Academy of Sciences of the United States of America. 107 Suppl 2 ع. Supplement 2: 8954–61. مايو 2010. Bibcode:2010PNAS..107.8954B. DOI:10.1073/pnas.0914618107. PMC:3024022. PMID:20445096.
- "Reconstructing the population history of Nicaragua by means of mtDNA, Y-chromosome STRs, and autosomal STR markers". American Journal of Physical Anthropology. ج. 143 ع. 4: 591–600. ديسمبر 2010. DOI:10.1002/ajpa.21355. PMID:20721944.
- "Genetic make up and structure of Colombian populations by means of uniparental and biparental DNA markers". American Journal of Physical Anthropology. ج. 143 ع. 1: 13–20. سبتمبر 2010. DOI:10.1002/ajpa.21270. PMID:20734436.
- "Analysis of Y chromosome SNPs in Alagoas, Northeastern Brazil". Forensic Science International: Genetics Supplement Series. ج. 2 ع. 1: 421–422. ديسمبر 2009. DOI:10.1016/j.fsigss.2009.08.166.
- "The Africa male lineages of Bahia's people—Northeast Brazil: A preliminary SNPs study". Forensic Science International: Genetics Supplement Series. ج. 2 ع. 1: 349–350. ديسمبر 2009. DOI:10.1016/j.fsigss.2009.07.010.
- Tanya M Simms 2011, The Peopling of the Bahamas: A Phylogeographical Perspective pg. 194
- "The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations". Annals of Human Genetics. ج. 65 ع. Pt 1: 43–62. يناير 2001. DOI:10.1046/j.1469-1809.2001.6510043.x. PMID:11415522.
- "Niger-Congo speaking populations and the formation of the Brazilian gene pool: mtDNA and Y-chromosome data". American Journal of Physical Anthropology. ج. 133 ع. 2: 854–67. يونيو 2007. DOI:10.1002/ajpa.20604. PMID:17427922.
- "The Levant versus the Horn of Africa: evidence for bidirectional corridors of human migrations". American Journal of Human Genetics. ج. 74 ع. 3: 532–44. مارس 2004. DOI:10.1086/382286. PMC:1182266. PMID:14973781.Luis JR, Rowold DJ, Regueiro M, Caeiro B, Cinnioğlu C, Roseman C, Underhill PA, Cavalli-Sforza LL, Herrera RJ (March 2004). "The Levant versus the Horn of Africa: evidence for bidirectional corridors of human migrations". American Journal of Human Genetics. 74 (3): 532–44. doi:10.1086/382286. PMC 1182266. PMID 14973781.
- "The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations". Annals of Human Genetics. ج. 65 ع. Pt 1: 43–62. يناير 2001. DOI:10.1046/j.1469-1809.2001.6510043.x. PMID:11415522.Underhill PA, Passarino G, Lin AA, Shen P, Mirazón Lahr M, Foley RA, Oefner PJ, Cavalli-Sforza LL (January 2001). "The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations". Annals of Human Genetics. 65 (Pt 1): 43–62. doi:10.1046/j.1469-1809.2001.6510043.x. PMID 11415522. S2CID 9441236.
- "Y-chromosomal variation in sub-Saharan Africa: insights into the history of Niger-Congo groups". Molecular Biology and Evolution. ج. 28 ع. 3: 1255–69. مارس 2011. DOI:10.1093/molbev/msq312. PMC:3561512. PMID:21109585.
- "The Bantu expansion revisited: a new analysis of Y chromosome variation in Central Western Africa". Molecular Ecology. ج. 20 ع. 13: 2693–708. يوليو 2011. DOI:10.1111/j.1365-294X.2011.05130.x. PMID:21627702.
- "Little genetic differentiation as assessed by uniparental markers in the presence of substantial language variation in peoples of the Cross River region of Nigeria". BMC Evolutionary Biology. ج. 10: 92. مارس 2010. DOI:10.1186/1471-2148-10-92. PMC:2867817. PMID:20356404.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - "Development of a single base extension method to resolve Y chromosome haplogroups in sub-Saharan African populations". Investigative Genetics. ج. 1 ع. 1: 6. سبتمبر 2010. DOI:10.1186/2041-2223-1-6. PMC:2988483. PMID:21092339.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - "Niger-Congo speaking populations and the formation of the Brazilian gene pool: mtDNA and Y-chromosome data". American Journal of Physical Anthropology. ج. 133 ع. 2: 854–67. يونيو 2007. DOI:10.1002/ajpa.20604. PMID:17427922.Hünemeier T, Carvalho C, Marrero AR, Salzano FM, Pena SD, Bortolini MC (June 2007). "Niger-Congo speaking populations and the formation of the Brazilian gene pool: mtDNA and Y-chromosome data". American Journal of Physical Anthropology. 133 (2): 854–67. doi:10.1002/ajpa.20604. PMID 17427922.
- "Sub-populations within the major European and African derived haplogroups R1b3 and E3a are differentiated by previously phylogenetically undefined Y-SNPs". Human Mutation. ج. 28 ع. 1: 97. يناير 2007. DOI:10.1002/humu.9469. PMID:17154278.Sims LM, Garvey D, Ballantyne J (January 2007). "Sub-populations within the major European and African derived haplogroups R1b3 and E3a are differentiated by previously phylogenetically undefined Y-SNPs". Human Mutation. 28 (1): 97. doi:10.1002/humu.9469. PMID 17154278. S2CID 34556775.
- "Little genetic differentiation as assessed by uniparental markers in the presence of substantial language variation in peoples of the Cross River region of Nigeria". BMC Evolutionary Biology. ج. 10: 92. مارس 2010. DOI:10.1186/1471-2148-10-92. PMC:2867817. PMID:20356404.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link)Veeramah KR, Connell BA, Ansari Pour N, Powell A, Plaster CA, Zeitlyn D, Mendell NR, Weale ME, Bradman N, Thomas MG (March 2010). "Little genetic differentiation as assessed by uniparental markers in the presence of substantial language variation in peoples of the Cross River region of Nigeria". BMC Evolutionary Biology. 10: 92. doi:10.1186/1471-2148-10-92. PMC 2867817. PMID 20356404. - "Ethiopians and Khoisan share the deepest clades of the human Y-chromosome phylogeny". American Journal of Human Genetics. ج. 70 ع. 1: 265–8. يناير 2002. DOI:10.1086/338306. PMC:384897. PMID:11719903.Semino O, Santachiara-Benerecetti AS, Falaschi F, Cavalli-Sforza LL, Underhill PA (January 2002). "Ethiopians and Khoisan share the deepest clades of the human Y-chromosome phylogeny". American Journal of Human Genetics. 70 (1): 265–8. doi:10.1086/338306. PMC 384897. PMID 11719903.
- "A back migration from Asia to sub-Saharan Africa is supported by high-resolution analysis of human Y-chromosome haplotypes". American Journal of Human Genetics. ج. 70 ع. 5: 1197–214. مايو 2002. DOI:10.1086/340257. PMC:447595. PMID:11910562.Cruciani F, Santolamazza P, Shen P, Macaulay V, Moral P, Olckers A, Modiano D, Holmes S, Destro-Bisol G, Coia V, Wallace DC, Oefner PJ, Torroni A, Cavalli-Sforza LL, Scozzari R, Underhill PA (May 2002). "A back migration from Asia to sub-Saharan Africa is supported by high-resolution analysis of human Y-chromosome haplotypes". American Journal of Human Genetics. 70 (5): 1197–214. doi:10.1086/340257. PMC 447595. PMID 11910562.
- "The Bantu expansion revisited: a new analysis of Y chromosome variation in Central Western Africa". Molecular Ecology. ج. 20 ع. 13: 2693–708. يوليو 2011. DOI:10.1111/j.1365-294X.2011.05130.x. PMID:21627702.Montano V, Ferri G, Marcari V, Batini C, Anyaele O, Destro-Bisol G, Comas D (July 2011). "The Bantu expansion revisited: a new analysis of Y chromosome variation in Central Western Africa". Molecular Ecology. 20 (13): 2693–708. doi:10.1111/j.1365-294X.2011.05130.x. PMID 21627702. S2CID 9951365.
- "Y-chromosomal variation in sub-Saharan Africa: insights into the history of Niger-Congo groups". Molecular Biology and Evolution. ج. 28 ع. 3: 1255–69. مارس 2011. DOI:10.1093/molbev/msq312. PMC:3561512. PMID:21109585.de Filippo C, Barbieri C, Whitten M, Mpoloka SW, Gunnarsdóttir ED, Bostoen K, Nyambe T, Beyer K, Schreiber H, de Knijff P, Luiselli D, Stoneking M, Pakendorf B (March 2011). "Y-chromosomal variation in sub-Saharan Africa: insights into the history of Niger-Congo groups". Molecular Biology and Evolution. 28 (3): 1255–69. doi:10.1093/molbev/msq312. PMC 3561512. PMID 21109585.
- "A map of human genome variation from population-scale sequencing". Nature. ج. 467 ع. 7319: 1061–73. أكتوبر 2010. Bibcode:2010Natur.467.1061T. DOI:10.1038/nature09534. PMC:3042601. PMID:20981092.
- Reynolds، David؛ Squecco، Adriano. "Y-Chromosome Genome Comparison". مؤرشف من الأصل في 2022-12-26. اطلع عليه بتاريخ 2011-08-01.
- "The imprint of the Slave Trade in an African American population: mitochondrial DNA, Y chromosome and HTLV-1 analysis in the Noir Marron of French Guiana". BMC Evolutionary Biology. ج. 10: 314. أكتوبر 2010. DOI:10.1186/1471-2148-10-314. PMC:2973943. PMID:20958967.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - "A map of human genome variation from population-scale sequencing". Nature. ج. 467 ع. 7319: 1061–73. أكتوبر 2010. Bibcode:2010Natur.467.1061T. DOI:10.1038/nature09534. PMC:3042601. PMID:20981092.Abecasis GR, Altshuler D, Auton A, Brooks LD, Durbin RM, Gibbs RA, Hurles ME, McVean GA (October 2010). "A map of human genome variation from population-scale sequencing". Nature. 467 (7319): 1061–73. Bibcode:2010Natur.467.1061T. doi:10.1038/nature09534. PMC 3042601. PMID 20981092.
- "Y-SNP analysis in an Angola population". Forensic Science International: Genetics Supplement Series. ج. 3 ع. 1: e369–e370. ديسمبر 2011. DOI:10.1016/j.fsigss.2011.09.046.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Y chromosome sequence variation and the history of human populations". Nature Genetics. ج. 26 ع. 3: 358–61. نوفمبر 2000. DOI:10.1038/81685. PMID:11062480.
- "Development of a single base extension method to resolve Y chromosome haplogroups in sub-Saharan African populations". Investigative Genetics. ج. 1 ع. 1: 6. سبتمبر 2010. DOI:10.1186/2041-2223-1-6. PMC:2988483. PMID:21092339.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link)Naidoo T, Schlebusch CM, Makkan H, Patel P, Mahabeer R, Erasmus JC, Soodyall H (September 2010). "Development of a single base extension method to resolve Y chromosome haplogroups in sub-Saharan African populations". Investigative Genetics. 1 (1): 6. doi:10.1186/2041-2223-1-6. PMC 2988483. PMID 21092339. - MacAulay، Vincent، المحرر (يناير 2011). "A new topology of the human Y chromosome haplogroup E1b1 (E-P2) revealed through the use of newly characterized binary polymorphisms". PLOS ONE. ج. 6 ع. 1: e16073. Bibcode:2011PLoSO...616073T. DOI:10.1371/journal.pone.0016073. PMC:3017091. PMID:21253605.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link)Trombetta B, Cruciani F, Sellitto D, Scozzari R (January 2011). MacAulay V (ed.). "A new topology of the human Y chromosome haplogroup E1b1 (E-P2) revealed through the use of newly characterized binary polymorphisms". PLOS ONE. 6 (1): e16073. Bibcode:2011PLoSO...616073T. doi:10.1371/journal.pone.0016073. PMC 3017091. PMID 21253605. - "New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree". Genome Research. ج. 18 ع. 5: 830–8. مايو 2008. DOI:10.1101/gr.7172008. PMC:2336805. PMID:18385274.
- All were positive for U175.
- The publication refers to E-V38 as H22.
- E-M2 is approximately 7.7–7.9% of total US male population.
- The publication transposes M116.2 with M116.1 in Table 1.
- The YCAII STR marker value of 19–19 is also usually indicative of U175.