المجموعة الفردانية E
المجموعة الفردانية E أو هابلوغروب E وتُسمى (E-M96) هي مجموعة هابلوغروب DNA لكروموسوم Y البشري.
| النمط الجماعي E | |
|---|---|
| الوقت المحتمل للنشوء | 65,200 - 69,000 - 73,000 سنة مضت |
| المكان المحتمل للنشوء | شرق إفريقيا، غرب إفريقيا، أوراسيا |
| عمر الالتحام | 52,300 سنة مضت |
| السلف | DE |
| السُّلان | E-P147 - E-M75 |
| الطفرة المعرّفة | L339, L614, M40/SRY4064/SRY8299, M96, P29, P150, P152, P154, P155, P156, P162, P168, P169, P170, P171, P172, P173, P174, P175, P176 |
وهو أحد الفرعين الرئيسيين للمجموعة الفردانية الأقدم والأسلف DE، والفرع الرئيسي الآخر هو المجموعة الفردانية D.
تنقسم المجموعة الحيوية E-M96 إلى فئتين فرعيتين رئيسيتين: الفئة الأكثر شيوعًا E-P147 والأقل شيوعًا E-M75.
الأصول
اقترح أندرهيل (2001) أن المجموعة الفردانية E ربما تكون قد نشأت في شرق أفريقيا.[1]
يقبل بعض المؤلفين مثل شاندراسيكار (2007)، الموقف السابق لهامر (1997) بأن المجموعة الفردانية E ربما تكون قد نشأت في غرب آسيا، [2] نظرًا لما يلي:
- E عبارة عن فرع حيوي من المجموعة الفردانية DE، بينما يتم توزيع الفرع الحيوي الرئيسي الآخر، المجموعة الفردانية D، حصريًا في آسيا.
- DE عبارة عن فرع حيوي داخل M168 مع الفرعين الرئيسيين الآخرين، C وF، اللذان يعتبران من أصل أوراسي بالفعل.
ومع ذلك، يُعتقد أن العديد من الاكتشافات التي تم إجراؤها منذ مقالات هامر تجعل الأصل الآسيوي أقل احتمالًا:
- أثبت أندرهيل وكيفيسيلد (2007) أن C وF لهما سلف مشترك مما يعني أن DE لديه شقيق واحد فقط وهو غير إفريقي.[3]
- تم العثور على DE في كل من آسيا وإفريقيا، مما يعني أنه لا يوجد واحد فقط، بل يوجد العديد من أشقاء D في آسيا وإفريقيا.
- قام كارافيت (2008)، والذي شارك هامر في تأليفه، بإعادة ترتيب التقديرات الزمنية بشكل كبير مما أدى إلى "تفسيرات جديدة حول الأصل الجغرافي للمجموعات الفرعية القديمة".[4]من بين أشياء أخرى، اقترحت هذه المقالة عمرًا أكبر بكثير للمجموعة الفردانية E-M96 مما تم اعتباره سابقًا، مما يمنحها عمرًا مشابهًا للمجموعة الفردانية D، وDE نفسها، مما يعني أنه لم يعد هناك أي سبب قوي لرؤيتها على أنها فرع من المجموعة الفردانية D. DE الذي يجب أن يكون قد ظهر بعد وقت طويل من ظهور DE إلى الوجود ودخوله إلى آسيا.[4]
كول وآخرون. (2009) افترض أن أصل المجموعة الفردانية E من غرب إفريقيا، قائلاً: "من بين المجموعات الفردانية العشرين الرئيسية في شجرة المجموعة الفردانية الكروموسومية Y، تم اكتشاف 5 فقط في السكان الأمهرية الذين تم تحليلهم في إثيوبيا.
المجموعة الفردانية A موجودة بالقرب من جذور الشجرة وتوجد فقط بين الذكور في القارة الأفريقية.
المجموعة الفردانية الرئيسية التي تم اكتشافها هي E. المجموعة الفردانية E لها أصلها في غرب إفريقيا.
وبسبب الهجرة فإن المجموعة الفردانية A التي كانت سائدة في الأصل في إثيوبيا قد تم استبدالها جزئيًا."
في عام 2015، بوزنيك وأندرهيل وآخرون. ادعى أن المجموعة الفردانية E نشأت خارج إفريقيا، بحجة أن "هذا النموذج من الفصل الجغرافي داخل الفرع الحيوي CT يتطلب فقط تبادل مجموعة هابلوغروبية قارية واحدة (E إلى إفريقيا)، بدلاً من ثلاثة (D، C، وF خارج إفريقيا).
إن توقيت هذه العودة المفترضة إلى إفريقيا، بين ظهور المجموعة الفردانية E وتمايزها داخل إفريقيا بمقدار 58 ألف سنة، يتوافق مع المقترحات المستندة على بيانات غير الكروموسوم Y، الخاصة بتدفق الجينات الوفير بين إفريقيا والجزيرة العربية بمقدار 50-80 ألف سنة."[5]
في عام 2015، تروبيتا وآخرون. اقترح أصل شرق إفريقي للمجموعة الفردانية E، قائلًا: "تحليلنا الجغرافي، استنادًا إلى آلاف العينات في جميع أنحاء العالم، يشير إلى أن إشعاع المجموعة الفردانية E بدأ حوالي 58 كا، في مكان ما في إفريقيا جنوب الصحراء الكبرى، مع احتمال خلفي أعلى (0.73). من أصل شرق إفريقي." [6]
كابريرا وآخرون. (2018) يفترض وجود مركز منشأ أوراسي وانتشار للمجموعة الفردانية E استنادًا إلى العمر المماثل للمجموعة الفردانية الأصلية للفرع الحيوي DE ومجموعة الفردانية mtDNA L3.
وفقًا لهذه الفرضية، بعد الهجرة الأولية خارج إفريقيا للبشر المعاصرين من الناحية التشريحية حوالي 125 ألف سنة، تنوعت المجموعة الفردانية DE حول جبال الهيمالايا وفي التبت أو غربها، وبعد ذلك يُقترح أن الذكور الحاملين للإلكترون قد هاجروا عائدين من مكان منشأ المجموعة الفردانية الأبوية في أوراسيا حوالي 70 ألف سنة، جنبًا إلى جنب مع الإناث اللاتي يحملن المجموعة الفردانية الأمومية L3، والتي تفترض الدراسة أيضًا أنها نشأت في أوراسيا، إلى إفريقيا.
ثم تم اقتراح أن هذه الأنساب الأوراسية الجديدة قد حلت إلى حد كبير محل الذكور الأصليين القديم (مثل المجموعة الفردانية B-M60) والأنساب الأفريقية الأنثوية.[7]
هابر وآخرون. (2019) اقترحت دراسة أصلًا إفريقيًا للمجموعة الفردانية E بناءً على تحليل البنية التطورية للكروموسومات Y، وأوقات تباعد المجموعة الفردانية، ومجموعة الفردانية D0 المكتشفة مؤخرًا والتي تم العثور عليها في ثلاثة نيجيريين، وهو فرع إضافي من سلالة DE يتباعد مبكرًا عن المجموعة الفردانية D يدعم المؤلفون الأصل الأفريقي للمجموعة الفردانية DE، وهجرة المجموعات الفردانية C وD و FT خارج أفريقيا قبل حوالي 50300-81000 سنة مضت.
تم العثور على تواريخ الاختلاف المبكرة في دراسة DE وE وD0 (جميعها مؤرخة بحوالي 71-76 ألف سنة)، والتي تم تحديدها لتسبق هجرة أسلاف الأوراسيين خارج أفريقيا (التي يعود تاريخها إلى حوالي 50-50 سنة). 60 كيا)، يعتبرها المؤلفون أيضًا أنها تدعم الأصل الأفريقي لتلك المجموعات الفردانية.
الحمض النووي القديم
وُجد أن بقايا العصر الحجري الحديث ما قبل الفخاري B من بلاد الشام كانت تحمل المجموعة الفردانية E (1/7؛ ~14%). [8]
في نياريندي روكشيلتر في كينيا، كان هناك شخصان يعود تاريخهما إلى العصر الحجري المتأخر (3500 سنة مضت)؛ أحدهما يحمل المجموعة الفردانية L4b2a والآخر يحمل المجموعة الفردانية E (E-M96، E-P162). [9] [10]
في كيندوكي في جمهورية الكونغو الديمقراطية، كان هناك ثلاثة أفراد، يعود تاريخهم إلى فترة ما قبل التاريخ (230 قبل الحاضر، 150 قبل الحاضر، 230 قبل الحاضر)؛ حملت إحداهما المجموعات الفردانية E1b1a1a1d1a2 (E-CTS99، E-CTS99) وL1c3a1b، وحملت أخرى المجموعة الفردانية E (E-M96، E-PF1620)، وحملت المجموعة الفردانية الأخيرة R1b1 (R-P25 1، R-M415) و L0a1b1a1.[11] [12]
توزيع الفروع
ينتمي معظم أعضاء المجموعة الفردانية E-M96 إلى E1، في حين أن المجموعة الفردانية E2-M75 نادرة.
وتنقسم المجموعة الفردانية E1a إلى فروع رئيسية: E1a1 (E-M44) والتي توجد في الغالب في أوروبا وغرب آسيا وبين اليهود الأشكناز؛ وE1a2 (E-Z958) الذي تم العثور عليه حصريًا في إفريقيا جنوب الصحراء الكبرى.[13]
المجموعة الفردانية E-M2 هي الفئة الفرعية الأكثر انتشارًا من الفئة E في إفريقيا جنوب الصحراء الكبرى وترتبط بقوة بتوسع متحدثي البانتو.
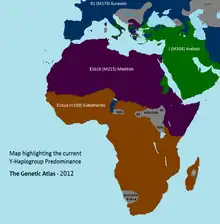
تم العثور على E-M215 على ترددات عالية في شمال أفريقيا وغرب آسيا وشرق أفريقيا وأوروبا.
E-M215 هو الأكثر شيوعًا بين المتحدثين باللغة الأفرو آسيوية في الشرق الأدنى وشمال أفريقيا والقرن الأفريقي، وقد تم العثور عليه أيضًا بين بعض المتحدثين باللغة النيلية الصحراوية والنيجر والكونغو في شمال شرق أفريقيا والسودان.
يعد E-M215 أقل شيوعًا بكثير في غرب ووسط وجنوب إفريقيا، على الرغم من أنه لوحظ بين بعض المتحدثين الخويسانيين[14] وبين المتحدثين بالنيجر والكونغو في سنيجامبيا،[15] غينيا بيساو،[16] بوركينا فاسو،[17] غانا، الجابون،[18] جمهورية الكونغو الديمقراطية، رواندا،[19] ناميبيا، وجنوب إفريقيا.
الفئات الفرعية
E-M96*
تشير Paragroup E-M96* إلى السلالات التي تنتمي إلى الفئة E ولكن لا يمكن تصنيفها في أي فرع معروف.
تم العثور على E(xE1-P147, E2-M75) - أي E الذي كان اختباره سلبيًا لكل من P147 وM75 - في 3 ذكور من لبنان،[20] 2 أمهرة من إثيوبيا،[21] ذكران من سوريا،[22] ذكران من المملكة العربية السعودية،[23] وذكر واحد يتحدث لغة البانتو من جنوب إفريقيا.[24]
تم العثور على E(xE1a-M33, E1b1-P2, E2-M75) بين العديد من سكان جنوب إفريقيا وفي رجل مصري؛[25] تمت ملاحظة E(xE1a-M33, E1b1a1-M2, E1b1b-M215, E2-M75) أيضًا بين الأقزام والبانتو من الكاميرون والجابون؛[26] وأيضا في بوركينا فاسو[27] ورجل فولبي من النيجر.[28]
تم اكتشاف مؤخرًا أن 3 رجال من شرق إفريقيا تم تصنيفهم سابقًا على أنهم E-M96* فقط يمكن تعيينهم لفرع جديد، E-V44، وهو فرع شقيق لـ E1-P147؛
يتشارك E-P147 وE-V44 في طفرة V3725، مما يجعل E2-M75 وE-V3725 الفرعين الأساسيين المعروفين لـ E.[29]
كما تم اكتشاف أن اثنين من المختبرين السعوديين الخاصين من مكة وجيزان ينتميان إلى هذا الفرع المراوغ والنادر.[30]
من غير المعروف ما إذا كانت بعض (أو كل) E * (xE1، E2) الأخرى في الدراسات السابقة ستندرج في V44 أيضًا أم لا.
E-P147
يعد E-P147 (المعروف أيضًا باسم E1) هو الفرع الأكثر عددًا والموزع على نطاق واسع من E-M96.
وله فرعين أساسيين: E-M132 (E1a) و E-P177 (E1b). تنقسم المجموعة الفردانية E1a إلى فرعين: E1a1 (E-M44) والتي تم العثور عليها في الغالب في أوروبا وغرب آسيا وبين اليهود الأشكناز؛ وE1a2 (Z958) الذي تم العثور عليه حصريًا في أفريقيا جنوب الصحراء الكبرى.[31]
المجموعة الفردانية E-P2 (E1b1) هي النوع الأكثر شيوعًا من E-M96 وسلالة Y-DNA الأكثر شيوعًا في إفريقيا مع سليلتين رئيسيتين: E-V38 (E1b1a) و E-M215 (E1b1b).
تم العثور عن المجموعة الفردانية E (xE3b,E3a) - أي أن اختبار E سلبي لكل من M35 وM2، تم العثور عليه في 11 ذكرًا من المغرب في Zalloua et al. (2008 ب). [22]
المجموعة الفردانية E-V38 هي سلف E-M2 (E1b1a1) وهي الفئة الفرعية الأكثر شيوعًا من E في مجمل إفريقيا جنوب الصحراء الكبرى، وترتبط بقوة بتوسع متحدثي البانتو في جميع أنحاء وسط وجنوب إفريقيا.
سليل آخر من E-V38، E-M329 (E1b1a2)، تمت ملاحظته في مجتمع صياد إثيوبي منذ 4200 سنة مضت، ويوجد في الغالب في الذكور من القرن الإفريقي وشبه الجزيرة العربية.[32]
من ناحية أخرى، يتم توزيع المجموعة الفردانية E-M215 (E1b1b) بترددات عالية في جميع أنحاء شمال إفريقيا وغرب آسيا وشرق إفريقيا وأوروبا.
تم العثور على المجموعة الفردانية E-Z827 في العينات النطوفية (E-Z830+) التي يرجع تاريخها إلى 10,000 سنة مضت من فلسطين، وتوجد بشكل شائع في جميع أنحاء غرب آسيا، وشمال إفريقيا، وأوروبا، وإثيوبيا.
تم العثور أيضًا على المجموعة الفردانية E-V68 بشكل شائع في شمال إفريقيا وغرب آسيا، وقد تم العثور عليها في البقايا الإيبيروموروسية التي يرجع تاريخها إلى 15000 سنة مضت من المغرب، مع سليلها الغزير E-V32 الذي يهيمن على سلالات الذكور في القرن الإفريقي.
E-M75
E-M75 (المعروف أيضًا باسم E2) موجود في جميع أنحاء إفريقيا شبه الاستوائية، وخاصة في منطقة البحيرات الكبرى الأفريقية ووسط أفريقيا.
تم العثور على أعلى تركيز للمجموعة الفردانية بين آلور (66.67%)،[33] هيما (38.89%)،[25] ريمايبي (27.03%)،[34] مبوتي (25.00%)،[17] دابا (22.22) %)،[17] إيفيا (20.83%)،[26] الزولو (20.69%)،[25] والبانتوس الكيني (17.24%).[35]
تم العثور على المجموعة الفردانية E-M75 (xM41,M54) في 6% (1/18) من داما من ناميبيا،[33] 4% (1/26) من غاندا من أوغندا،[25] 3% (1/39) من الماندينكا من غامبيا/السنغال،[25] و2% (1/49) من الشونا من زيمبابوي. [25]
يُظهر اختبار الحمض النووي التجاري الخاص في Family Tree DNA العديد من ذكور E-M75 القادمين من شبه الجزيرة العربية (المملكة العربية السعودية والبحرين والكويت واليمن والإمارات العربية المتحدة)، وبين اليهود الأشكناز.[36]
علم الوراثة
التاريخ التطوري
قبل عام 2002، كان هناك في الأدبيات الأكاديمية ما لا يقل عن سبعة أنظمة تسمية لشجرة النشوء والتطور للكروموسوم Y.
وقد أدى هذا إلى ارتباك كبير.
في عام 2002، اجتمعت المجموعات البحثية الرئيسية معًا وشكلت اتحاد كروموسوم Y (YCC).
لقد نشروا ورقة مشتركة أنشأت شجرة واحدة جديدة اتفق الجميع على استخدامها.
وفي وقت لاحق، شكلت مجموعة من العلماء المواطنين المهتمين بعلم الوراثة السكانية وعلم الأنساب الجيني مجموعة عمل لإنشاء شجرة هواة تهدف قبل كل شيء إلى أن تكون في الوقت المناسب.
مراجع
- Underhill، P. A.؛ Passarino، G.؛ Lin، A. A.؛ Shen، P.؛ Mirazon Lahr، M.؛ Foley، R. A.؛ Oefner، P. J.؛ Cavalli-Sforza، L. L. (2001). "The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations". Annals of Human Genetics. ج. 65 ع. Pt 1: 43–62. DOI:10.1046/j.1469-1809.2001.6510043.x. PMID:11415522.
- Chandrasekar؛ Saheb، S. Y.؛ Gangopadyaya، P.؛ Gangopadyaya، S.؛ Mukherjee، A.؛ Basu، D.؛ Lakshmi، G. R.؛ Sahani، A. K.؛ Das، B.؛ وآخرون (سبتمبر–أكتوبر 2007). "YAP insertion signature in South Asia". Annals of Human Biology. ج. 34 ع. 5: 582–6. DOI:10.1080/03014460701556262. PMID:17786594.
- Underhill، Peter A.؛ Kivisild، Toomas (2007). "Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations". Annual Review of Genetics. ج. 41: 539–64. DOI:10.1146/annurev.genet.41.110306.130407. PMID:18076332.
- Karafet، T. M.؛ Mendez، F. L.؛ Meilerman، M. B.؛ Underhill، P. A.؛ Zegura، S. L.؛ Hammer، M. F. (2008). "New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree". Genome Research. ج. 18 ع. 5: 830–8. DOI:10.1101/gr.7172008. PMC:2336805. PMID:18385274.
- Poznik، G David؛ وآخرون (2016). "Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences". Nature Genetics. ج. 48 ع. 6: 593–599. DOI:10.1038/ng.3559. PMC:4884158. PMID:27111036.
- Trombetta et al. 2015, Phylogeographic refinement and large scale genotyping of human Y chromosome haplogroup E provide new insights into the dispersal of early pastoralists in the African continent
- Cabrera، Vicente؛ Marrero، Patricia؛ Abu-Amero، Khaled؛ Larruga، Jose (2019). "Carriers of mitochondrial DNA macrohaplogroup L3 basal lineages migrated back to Africa from Asia around 70,000 years ago". BMC Evolutionary Biology. ج. 18 ع. 98: 98. DOI:10.1186/s12862-018-1211-4. PMC:6009813. PMID:29921229.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - Martiniano، Rui؛ Sanctis، Bianca De؛ Hallast، Pille؛ Durbin، Richard (20 ديسمبر 2020). "Supplementary Material: Placing ancient DNA sequences into reference phylogenies". Molecular Biology and Evolution. ج. 39 ع. 2: 2020.12.19.423614. DOI:10.1093/molbev/msac017. PMC:8857924. PMID:35084493.
- Wang، Ke؛ وآخرون (2020). "Ancient genomes reveal complex patterns of population movement, interaction, and replacement in sub-Saharan Africa". Science Advances. ج. 6 ع. 24: eaaz0183. Bibcode:2020SciA....6..183W. DOI:10.1126/sciadv.aaz0183. ISSN:2375-2548. OCLC:8616876709. PMC:7292641. PMID:32582847.
- Wang، Ke؛ وآخرون (2020). "Supplementary Materials for Ancient genomes reveal complex patterns of population movement, interaction, and replacement in sub-Saharan Africa" (PDF). Science Advances. ج. 6 ع. 24: eaaz0183. Bibcode:2020SciA....6..183W. DOI:10.1126/sciadv.aaz0183. ISSN:2375-2548. OCLC:8616876709. PMC:7292641. PMID:32582847. مؤرشف من الأصل (PDF) في 2023-03-14.
- Wang، Ke؛ وآخرون (2020). "Ancient genomes reveal complex patterns of population movement, interaction, and replacement in sub-Saharan Africa". Science Advances. ج. 6 ع. 24: eaaz0183. Bibcode:2020SciA....6..183W. DOI:10.1126/sciadv.aaz0183. ISSN:2375-2548. OCLC:8616876709. PMC:7292641. PMID:32582847.Wang, Ke; et al. (2020). "Ancient genomes reveal complex patterns of population movement, interaction, and replacement in sub-Saharan Africa". Science Advances. 6 (24): eaaz0183. Bibcode:2020SciA....6..183W. doi:10.1126/sciadv.aaz0183. ISSN 2375-2548. OCLC 8616876709. PMC 7292641. PMID 32582847. S2CID 219604401.
- Wang، Ke؛ وآخرون (2020). "Supplementary Materials for Ancient genomes reveal complex patterns of population movement, interaction, and replacement in sub-Saharan Africa" (PDF). Science Advances. ج. 6 ع. 24: eaaz0183. Bibcode:2020SciA....6..183W. DOI:10.1126/sciadv.aaz0183. ISSN:2375-2548. OCLC:8616876709. PMC:7292641. PMID:32582847. مؤرشف من الأصل (PDF) في 2023-03-14.Wang, Ke; et al. (2020). "Supplementary Materials for Ancient genomes reveal complex patterns of population movement, interaction, and replacement in sub-Saharan Africa" (PDF). Science Advances. 6 (24): eaaz0183. Bibcode:2020SciA....6..183W. doi:10.1126/sciadv.aaz0183. ISSN 2375-2548. OCLC 8616876709. PMC 7292641. PMID 32582847. S2CID 219604401.
- E1a-M132 (Yfull) نسخة محفوظة 2023-07-28 على موقع واي باك مشين.
- Cruciani، Fulvio؛ La Fratta، Roberta؛ Santolamazza، Piero؛ Sellitto، Daniele؛ Pascone، Roberto؛ Moral، Pedro؛ Watson، Elizabeth؛ Guida، Valentina؛ وآخرون (2004). "Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out of Africa". The American Journal of Human Genetics. ج. 74 ع. 5: 1014–22. DOI:10.1086/386294. PMC:1181964. PMID:15042509.
- Wood، Elizabeth T؛ Stover، Daryn A؛ Ehret، Christopher؛ Destro-Bisol، Giovanni؛ Spedini، Gabriella؛ McLeod، Howard؛ Louie، Leslie؛ Bamshad، Mike؛ وآخرون (2005). "Contrasting patterns of Y chromosome and mtDNA variation in Africa: Evidence for sex-biased demographic processes". European Journal of Human Genetics. ج. 13 ع. 7: 867–76. DOI:10.1038/sj.ejhg.5201408. PMID:15856073. (cf. Appendix A: Y Chromosome Haplotype Frequencies)
- Rosa، Alexandra؛ Ornelas، Carolina؛ Jobling، Mark A؛ Brehm، António؛ Villems، Richard (2007). "Y-chromosomal diversity in the population of Guinea-Bissau: A multiethnic perspective". BMC Evolutionary Biology. ج. 7: 124. DOI:10.1186/1471-2148-7-124. PMC:1976131. PMID:17662131.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - Cruciani، Fulvio؛ Santolamazza، Piero؛ Shen، Peidong؛ MacAulay، Vincent؛ Moral، Pedro؛ Olckers، Antonel؛ Modiano، David؛ Holmes، Susan؛ وآخرون (2002). "A Back Migration from Asia to Sub-Saharan Africa is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes". The American Journal of Human Genetics. ج. 70 ع. 5: 1197–214. DOI:10.1086/340257. PMC:447595. PMID:11910562.
- Berniell-Lee، G.؛ Calafell، F.؛ Bosch، E.؛ Heyer، E.؛ Sica، L.؛ Mouguiama-Daouda، P.؛ Van Der Veen، L.؛ Hombert، J.-M.؛ وآخرون (2009). "Genetic and Demographic Implications of the Bantu Expansion: Insights from Human Paternal Lineages". Molecular Biology and Evolution. ج. 26 ع. 7: 1581–9. DOI:10.1093/molbev/msp069. PMID:19369595.
- Luis، J؛ Rowold، D؛ Regueiro، M؛ Caeiro، B؛ Cinnioglu، C؛ Roseman، C؛ Underhill، P؛ Cavallisforza، L؛ Herrera، R (2004). "The Levant versus the Horn of Africa: Evidence for Bidirectional Corridors of Human Migrations". The American Journal of Human Genetics. ج. 74 ع. 3: 532–44. DOI:10.1086/382286. PMC:1182266. PMID:14973781.
- Zalloua، Pierre (2008). "Y-Chromosomal Diversity in Lebanon Is Structured by Recent Historical Events". Am J Hum Genet. ج. 82 ع. 4: 873–882. DOI:10.1016/j.ajhg.2008.01.020. PMC:2427286. PMID:18374297.
- Abu-Amero، Khaled K؛ Hellani، Ali؛ González، Ana M؛ Larruga، Jose M؛ Cabrera، Vicente M؛ Underhill، Peter A (2011). "Variation in Y chromosome, mitochondrial DNA and labels of identity on Ethiopia". UCL Discovery. ج. 10: 59. DOI:10.1186/1471-2156-10-59. PMC:2759955. PMID:19772609.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - Zalloua، Pierre (2008). "Identifying Genetic Traces of Historical Expansions: Phoenician Footprints in the Mediterranean". Am J Hum Genet. ج. 83 ع. 5: 633–642. DOI:10.1016/j.ajhg.2008.10.012. PMC:2668035. PMID:18976729.
- Abu-Amero، Khaled K؛ Hellani، Ali؛ González، Ana M؛ Larruga، Jose M؛ Cabrera، Vicente M؛ Underhill، Peter A (2009). "Saudi Arabian Y-Chromosome diversity and its relationship with nearby regions". BMC Genetics. ج. 10: 59. DOI:10.1186/1471-2156-10-59. PMC:2759955. PMID:19772609.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - Karafet، T. M.؛ Mendez، F. L.؛ Meilerman، M. B.؛ Underhill، P. A.؛ Zegura، S. L.؛ Hammer، M. F. (2008). "New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree". Genome Research. ج. 18 ع. 5: 830–8. DOI:10.1101/gr.7172008. PMC:2336805. PMID:18385274.Karafet, T. M.; Mendez, F. L.; Meilerman, M. B.; Underhill, P. A.; Zegura, S. L.; Hammer, M. F. (2008). "New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree". Genome Research. 18 (5): 830–8. doi:10.1101/gr.7172008. PMC 2336805. PMID 18385274.
- Wood (2005). "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes". Eur J Hum Genet. ج. 13 ع. 7: 867–76. DOI:10.1038/sj.ejhg.5201408. PMID:15856073.
- Berniell-Lee، G.؛ Calafell، F.؛ Bosch، E.؛ Heyer، E.؛ Sica، L.؛ Mouguiama-Daouda، P.؛ Van Der Veen، L.؛ Hombert، J.-M.؛ وآخرون (2009). "Genetic and Demographic Implications of the Bantu Expansion: Insights from Human Paternal Lineages". Molecular Biology and Evolution. ج. 26 ع. 7: 1581–9. DOI:10.1093/molbev/msp069. PMID:19369595.Berniell-Lee, G.; Calafell, F.; Bosch, E.; Heyer, E.; Sica, L.; Mouguiama-Daouda, P.; Van Der Veen, L.; Hombert, J.-M.; et al. (2009). "Genetic and Demographic Implications of the Bantu Expansion: Insights from Human Paternal Lineages". Molecular Biology and Evolution. 26 (7): 1581–9. doi:10.1093/molbev/msp069. PMID 19369595.
- de Filippo (2011). "Y-chromosomal variation in Sub-Saharan Africa: insights into the history of Niger-Congo groups". Mol Biol Evol. ج. 28 ع. 3: 1255–1269. DOI:10.1093/molbev/msq312. PMC:3561512. PMID:21109585.
- Bučkova (2013). "Multiple and differentiated contributions to the male gene pool of pastoral and farmer populations of the African Sahel". Am J Phys Anthropol. ج. 151 ع. 1: 10–21. DOI:10.1002/ajpa.22236. PMID:23460272.
- Trombetta (2015). "Phylogeographic refinement and large scale genotyping of human Y chromosome haplogroup E provide new insights into the dispersal of early pastoralists in the African continent". Genome Biol Evol. ج. 7 ع. 7: 1940–50. DOI:10.1093/gbe/evv118. PMC:4524485. PMID:26108492.
- E-FTA45776 نسخة محفوظة 2023-07-28 على موقع واي باك مشين.
- E1a-M132 نسخة محفوظة 2023-07-28 على موقع واي باك مشين.
- E-M329 نسخة محفوظة 2023-04-11 على موقع واي باك مشين.
- Wood (2005). "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes". Eur J Hum Genet. ج. 13 ع. 7: 867–76. DOI:10.1038/sj.ejhg.5201408. PMID:15856073.Wood (2005). "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes". Eur J Hum Genet. 13 (7): 867–76. doi:10.1038/sj.ejhg.5201408. PMID 15856073.
- Cruciani، Fulvio؛ Santolamazza، Piero؛ Shen، Peidong؛ MacAulay، Vincent؛ Moral، Pedro؛ Olckers، Antonel؛ Modiano، David؛ Holmes، Susan؛ وآخرون (2002). "A Back Migration from Asia to Sub-Saharan Africa is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes". The American Journal of Human Genetics. ج. 70 ع. 5: 1197–214. DOI:10.1086/340257. PMC:447595. PMID:11910562.Cruciani, Fulvio; Santolamazza, Piero; Shen, Peidong; MacAulay, Vincent; Moral, Pedro; Olckers, Antonel; Modiano, David; Holmes, Susan; et al. (2002). "A Back Migration from Asia to Sub-Saharan Africa is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes". The American Journal of Human Genetics. 70 (5): 1197–214. doi:10.1086/340257. PMC 447595. PMID 11910562.
- Luis، J؛ Rowold، D؛ Regueiro، M؛ Caeiro، B؛ Cinnioglu، C؛ Roseman، C؛ Underhill، P؛ Cavallisforza، L؛ Herrera، R (2004). "The Levant versus the Horn of Africa: Evidence for Bidirectional Corridors of Human Migrations". The American Journal of Human Genetics. ج. 74 ع. 3: 532–44. DOI:10.1086/382286. PMC:1182266. PMID:14973781.Luis, J; Rowold, D; Regueiro, M; Caeiro, B; Cinnioglu, C; Roseman, C; Underhill, P; Cavallisforza, L; Herrera, R (2004). "The Levant versus the Horn of Africa: Evidence for Bidirectional Corridors of Human Migrations". The American Journal of Human Genetics. 74 (3): 532–44. doi:10.1086/382286. PMC 1182266. PMID 14973781.
- Family Tree DNA public haplotree, Haplogroup E-M75 نسخة محفوظة 2023-07-28 على موقع واي باك مشين.
- Platt, D.E., Artinian, H., Mouzaya, F. et al. Autosomal genetics and Y-chromosome haplogroup L1b-M317 reveal Mount Lebanon Maronites as a persistently non-emigrating population. Eur J Hum Genet 29, 581–592 (2021). 10.1038/s41431-020-00765-x نسخة محفوظة 2023-07-28 على موقع واي باك مشين.
 بوابة علم الإنسان
بوابة علم الإنسان بوابة ما قبل التاريخ
بوابة ما قبل التاريخ