أكروكانثوصور
الأكروكانثوصور (الاسم العلمي: Acrocanthosaurus)، (من الإغريقية:akra = ακρα = عالي + akantha = ακανθα =أشواك أو فقرات + sauros = σαυρος = سحلية)، وهو جنس من ديناصورات الكاركارودونتوصوريدات التي كانت موجودة في ما يعرف الآن بأمريكا الشمالية خلال مرحلة الأبتي وبداية مرحلة الألبي من الطباشيري المبكر. ومثل معظم أجناس الديناصورات، فإن جنس الأكروكانثوصور يشمل على نوع واحد فقط وهو (الأكروكانثوصور أتوكينسيس). تم العثور على بقاياها الأحفورية بشكل رئيسي في الولايات الأمريكية وتحديداً في ولايات أوكلاهوما وتكساس ووايومنغ. حتى أنه تم العثور على أسنان نسبت إلى الأكروكانثوصور في أقصى الشرق مثل ولاية ماريلند، وهذا يشير إلى وجود مجموعة منشرة من القارة.
اضغط هنا للاطلاع على كيفية قراءة التصنيف العصر: الطباشيري المبكر, 116–110 مليون سنة | |
|---|---|
 جمجمة أكروكونثوصور متحف كارولاينا الشمالية للعلوم الطبيعية. | |
| المرتبة التصنيفية | جنس |
| التصنيف العلمي | |
| المملكة: | حيوانات |
| الشعبة: | حبليات |
| الطائفة: | زواحف |
| الرتبة العليا: | ديناصور |
| الرتبة: | سحليات الورك |
| الرتيبة: | ثيروبودا |
| الفصيلة العليا: | قريبات الألوصورات |
| الفصيلة: | ألوصوريات or عظايا الكركردون |
| الجنس: | Acrocanthosaurus جاي ويليس ستوفال & Langston, 1950 |
| الاسم العلمي | |
| Acrocanthosaurus جون ويليس ستوفال و Wann Langston ، 1950 | |
| نوع | |
| |
| مرادفات | |
| |
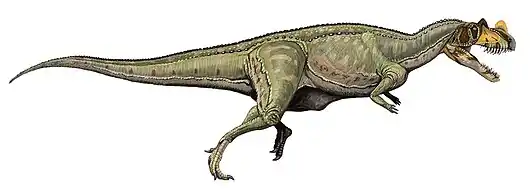
الأكروكانثوصور من الحيوانات المفترسة ثنائيات الحركة أي تمشي على قدمين فقط. وحسب اسمه المقترح فإنه يشتهر بارتفاع عدة فقرات من عموده الفقري العصبي، والتي على الأرجح تدعم السلسلة العضلية فوق رقبة الحيوان، وكذلك الظهر والوركين.[1] يعد الأكروكانثوصور أحد أكبر ديناصورات وحشيات الأرجل، حيث يصل طوله إلى 11.5 مترا، ويصل وزنه إلى 6.2 طن متري.[2] قد تكون آثار الأقدام لوحشيات الأرجل الكبيرة المكتشفة في تكساس قد كانت للكروكانثوصور، بالرغم من عدم وجود ارتباط مباشر ببقايا الهيكل العظمي.
أوضحت الاكتشافات الحديثة الكثير من التفاصيل في تشريح الأكروكانثوصور، مما سمح بالدراسات المتخصصة بأن تركز على بنية الدماغ ووظيفة الأطراف الأمامية. كان الأكروكانثوصور أكبر ديناصورات وحشيات الأرجل في نظامه البيئي ومن المحتمل أنه مفترس علوي يفترس سحليات الأرجل، وطيريات الأرجل والأنكيلوصوريات.
الاكتشاف والتسمية
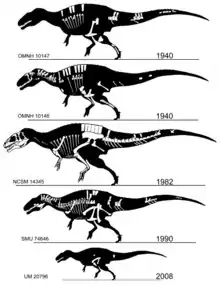
كما ذكر من قبل فقد سمي الأكروكانثوصور بسبب العمود الفقري العصبي الطويل، وقد جاءت (من الإغريقية:akra = ακρα = عالي + akantha = ακανθα =أشواك أو فقرات + sauros = σαυρος = سحلية).[3] وهناك نوع واحد فقط يسمى (الأكروكانثوصور أتوكينسيس) (الاسم العلمي: Acrocanthosaurus atokensis)، والمسمى على مقاطعة أتوكا في ولاية أوكلاهوما الأمريكية، التي وجدت فيه العينات الأصلية الخاصة به. وقد تمت صياغة هذا الاسم في عام 1950 من قبل عالمي الأحافير الأمريكيين «جون ويليس ستوفال» و«وان لانغستون جونيور».[1] اقترح «لانغستون» اسم (الأكراكانثوس أتوكينسيس) (الاسم العلمي: Acracanthus atokaensis) للجنس والأنواع في رسالته للماجستير غير المنشورة لعام 1947،[4][5] ولكن تم تغيير الاسم إلى (الأكروكانثوصور أتوكينسيس) للنشر الرسمي.
في بداية العقد 1940 تم اكتشاف النموذج النوعي والنموذج المجاور (OMNH 10146 و OMNH 10147)، وتم وصفهما في نفس الوقت في عام 1950، ويتكونان من جزئين لهيكلين وقطعة من مادة الجمجمة مستخرجة من تكوين أنتلرز في أوكلاهوما.[1] وفي العقد 1990 تم وصف عينتين أكثر اكتمالا، الأولى (SMU 74646) وهي عبارة عن جزء من هيكل عظمي، وقد تم تجميع أغلب الأجزاء المفقودة للجمجمة من تكوين جبال التوأم في تكساس وهي حاليا جزء من مجموعة متحف فورت وورث للعلوم والتاريخ.[6] وتم تجميع هيكل عظمي أكثر اكتمالا (NCSM 14345، الملقب بـ«فران») من تكوين انتلرز في أوكلاهوما بواسطة كل من «سيفيس هل» و«سايد لوف»، بترتيب من معهد بلاك هيلز في ولاية داكوتا الجنوبية، وهو موجود الآن في متحف نورث كارولينا للعلوم الطبيعية في رالي. وتعتبر هذه العينة هي الأكبر وتشمل الجمجمة الكاملة والأطراف الأمامية الوحيدة المعروفة.[7] وتعد العناصر الهيكلية لـ (OMNH 10147) بنفس الحجم تقريبا للعظام المماثلة في (NCSM 14345)، مما يشير إلى أن الحيوان بنفس الحجم تقريبا بينما النموذج النوعي و (SMU 74646) أصغر بكثير.[7]
تم توثيق وجود الأكروكانثوصور في تكوين كلوفري في عام 2012 بوصف هيكل عظمي جزئي آخر (UM 20796). تتكون هذه العينة من عدة أجزاء التي تمثل حيوانا صغيرا، مثل الفقرتين، وعظام عانة جزئية، وعظم الفخذ، وعظم شظية جزئي وبعض الفتات. وقد جائت هذه العينات من طبقة عظمية من حوض بيغورن في شمال وسط وايومنغ، ووقد عثر عليها بالقرب من عظم كتف ديناصور الصوروبوسيدون. قد تنتمي مجموعة متنوعة أخرى لبقايا مجزأة من وحشيات الأرجل في نفس التكوين إلى الأكروكانثوصور، والتي قد تكون وحشيات الأرجل الكبير الوحيدة في تكوين كلوفري.[8]
الوصف

كان الأكروكانثوصور من بين أكبر وحشيات الأرجل المعروفة في وقته. وتشير التقديرات إلى أن أكبر عينة معروفة (NCSM 14345) قد بلغت 11.5 مترا[2] من الخطم إلى طرف الذيل وتزن من 5.7 إلى 6.2 طن،[2][9] وأقصى وزن بلغ 7.25 طن ضمن نطاق إمكانية هذه العينة. ويقدر طول جمجمتها وحدها حوالي 1.3 متر.[7]
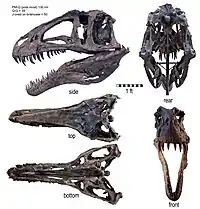
تشبه جمجمة الأكروكانثوصور معظم الألوصورات فهي طويلة ومنخفضة ونحيلة. وكانت ثقوب أمام العينية الخافضة للوزن كبيرة جدا، فقد كانت أكثر من ربع طول الجمجمة وثلثي ارتفاعها. لم يكن السطح الخارجي للفك العلوي والسطح العلوي لعظم الأنف لسطح الخطم خشنا كما في الجيجانوتوصور أو الكاركارودونتوصور. وقد نشأت نتوءات طويلة ومنخفضة من عظام الأنف، مستمرة على علي جانبي الخطم من فتحة الأنف وحتى العين، مستمرة حتى العظام الدمعية.[7] وهذه هي السمة الخاصة لجميع الألوصورات.[10] وعلى عكس الألوصور لم يكن هناك قزعة بارزة على العظم الدمعي أمام العين. اجتمعت العظام الدمعية والخَلفمَحجَريّة لتشكل حاجبا كثيفا فوق العين، كما رأينا في الكاركارودونتوصوريدات والأبليصوريدات الغير متعلقة. لديها تسعة عشر سنا منحني ومشرشرة تصطف على جانبي الفك العلوي، ولم يعرف عدد الأسنان للفك السفلي. وقد كانت أسنان الأكروكانثوصور أعرض من أسنان الكاركارودونتوصورات التي لم يكن لديها نسيج مجعد يميز الكاركارودونتوصوريدات. يتميز العظم السني (عظم الفك السفلي الحامل للأسنان) بأنه مربع الشكل عند الحافة الأمامية كما في الجيجانوتوصور وسطحية، بينما باقي الفك يكون خلفه وعميق جدا. يشترك الأكروكانثوصور والجيجانوتوصور في سلسلة من النتوءات الأفقية السميكة على السطح الخارجي لعظم فوق الزاوية للفك السفلي، تحت المفصل مع الجمجمة.[7]
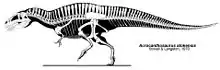
أبرز ما يميز الأكروكانثوصور هي الأشواك الظهرية الطويلة التي تقع على فقرات العنق والظهر والوركين وأعلى الذيل، والتي يمكن أن يزيد ارتفاعها عن 2.5 ضعف ارتفاع الفقرات التي امتدت منها.[1] تمتلك الديناصورات الأخرى أيضًا أشواك عالية على ظهرها، وفي بعض الأحيان أعلى بكثير من تلك الموجودة في الأكروكانثوصور. على سبيل المثال، الجنس الأفريقي السبينوصور لديه أشواك بطول مترين تقريبا، أطول بحوالي 11 مرة من فقراتها.[11] يرتبط بالأشواك السفلية للأكروكانثوصور عضلات قوية كما في البيسون الحديث، ربما تشكل ضلوع طويلة وسميكة أسفل ظهرها.[1] لا تزال وظيفة هذه الأشواك غير معروفة، ربما تكون للتواصل، أو لتخزين الدهون، أو للتحكم في العضلات أو درجة الحرارة. كل الفقرات العنقية والظهرية (الخلفية) بها أغواط محدبة على الجانبين، بينما تكون أصغر في الفقرات الذيلية. وهذا سبيه بالكاركارودونتوصوريدات أكثر من الألوصورات.[6]
بصرف النظر عن فقراته فقد كان للأكروكانثوصور هيكل نموذجي للألوصورات. ويعتبر الأكروكانثوصور من ذوات القَدَمَيْن وله ذيل طويل ثقيل يوازن الرأس والجسم، ليحافظ على مركز جاذبيته على الوركين. وأطرافه الأمامية أقصر وأكثر قوة من التي لدى الألوصور ولكنها متشابهة حيث أن كل يد تحمل ثلاثة أصابع بمخالب. لم يكن الأكروكانثوصور سريعا في الجري على عكس العديد من الديناصورات الصغيرة السريعة،[6][7] لأن عظم الفخذ أطول من عظم الساق والمشط.[12] وما لا يثير الدهشة هو أن عظام الساق الخلفية للأكروكانثوصور أكثر قوة من عظام أقربائها اللوصورات الأصغر. في كل قدم من أربعة أصابع رغم أن هذا هو الحال بالنسبة لوحشيات الأرجل، فالأول منها كان أصغر بكثير من البقية ولا يلامس الأرض.[6][7]
التصنيف والنظاميات
يصنف الأكروكانثوصور في الفصيلة العليا قريبات الألوصورات ضمن دون الرتبة صلبات الذيل. تتميز هذه الفصيلة العليا بوجود نتوءات مزدوجة على عظام الأنف والدمعية أعلى الأنف وتوجد أشواك ظهرية طويلة على فقرات العنق، ولها ميزات أخرى.[10] وقد تم وضعه في الأصل في فصيلة الألوصوريات مع الألوصور،[1] وهذا الترتيب مدعوم كذلك بدراسات تمت حتى أواخر عام 2000.[7] وقد وجدت معظم الدراسات أنه ينتمي إلى الفصيلة المرتبطة الكاركارودونتوصوريدات.[10][13][14]
قد عرف الأكروكانثوصور عند اكتشافه مع معظم وحشيات الأرجل الكبيرة الأخرى من بقايا مجزأة فقط، وقد أدى هذا إلى تصنيفات متغايرة جدا لهذا الجنس. قام «جون ويليس ستوفال» و«وان لانغستون جونيور» تم ربطها لأول مرة مع "Antrodemidae" المرادفة للألوصوريات، ولكن في عام 1956 تم نقلها بواسطة عالم الحيوانات ألفرد رومر إلى أصنوفة سلة مهملات الميجالوصورات.[15] وبعض العلماء يعتقد إلى أن له علاقة مع السبينوصور بسبب وجود الأشواك الطويلة على عموده الفقري.[16][17] استمر هذا الاعتقاد باعتبار الأكروكانثوصور من السبينوصوريات حتى العقد 1980،[18] وأنتشرت في كتب الديناصورات شبه التقنية في ذلك الوقت.[19][20]
كانت جميع ذوات الفقرات الشوكية الطويلة التي من أوائل العصر الطباشيري في إنجلترا تعتبر مشابهة جدا لتلك الموجودة في الأكروكانثوصور،[21] وفي عام 1988 اسماها العالم «غريغوري باول» كنوع ثاني من الجنس «الأكروكانثوصور ألتيسبيناكس» (الاسم العلمي: A. altispinax).[22] وقد ربطت هذه العظام بالأصل لديناصور الألتيسبيناكس المعروف من وحشيات الأرجل الإنجليزي والذي عرف فقط من الأسنان، وأدى هذا الربط إلى اقتراح عالم واحد على الأقل بأن الألتيسبيناكس نفسه كان مرادفا للأكروكانثوصور.[21] فيما بعد تم ربط هذه الفقرات للجنس الجديد «باكلسبيناكس» (الاسم العلمي: Becklespinax)، المنفصل نهائيا عن كل من الأكروكانثوصور والألتيسبيناكس.[23]

بينت معظم تحليلات التصنيف التفرعي التي تشمل الأكروكانثوصور بأنها كاركارودونتوصوريدات، وعادة في وضع التصنيف قاعدي المتعلق بالكاركارودونتوصور الأفريقي والجيجانوتوصور من أمريكا الجنوبية.[6][10][24] وغالبا ما كان يعتبر بأنه الأصنوفة الشقيقة لديناصور الإيوكارشاريا القاعدية من أفريقيا أيضا. وغالبا ما يُعتبر النيوفيناتور المكتشف في إنجلترا من الكاركارودونتوصوريدات الأكثر قاعدية أو كعضو قاعدي في مجموعة شقيقة تسمى النيوفيناتوريات.[12][14] وهذا يشير إلى أن الفصائل قد نشأت في أوروبا ثم انتشرت في القارات الجنوبية (في ذلك الوقت كانت متحدة باسم القارة العملاقة غندوانا). إذا كان الأكروكانثوصور من الكاركارودونتوصوريدات، إذا من الممكن أن يحدث الانتشار كذلك في أمريكا الشمالية.[6] عاشت جميع الكاركارودونتوصوريدات المعروفة خلال الفترة من بداية العصر الطباشيري وحتى منتصفه.[10]
يُظهر مخطط النسل التالي الذي أعده «نوفاس» مع بعض العلماء الأخرين في 2013 موضع الأكروكانثوصور داخل الكاركارودونتوصوريدات:[25]
| |||||||||||||||||||||||||||||||||||||||||||||||||
البيولوجيا القديمة
النمو والعمر

تشير التقديرات من السمات العظمية للنموذجين النوعيين "OMNH 10146" و "NCSM 14345" إلى أن الأكروكانثوصور يحتاج إلى 12 عاما على الأقل لينمو بشكل كامل. وقد يكون هذا الرقم أعلى من ذلك بكثير لأنه خلال عملية إعادة بناء العظام ونمو التجويف النخاعي فقدت بعض خطوط توقف النمو (خطوط هاريس). وإذا حسبنا هذه الخطوط فإن الأكروكانثوصور يحتاج ما بين 18-24 سنة حتى ينضج.[26]
وظيفة الأطراف الأمامية
مثل معظم وحشيات الأرجل الغير طيرية الأخرى فإن أطراف الأكروكانثوصور الأمامية لا تلامس الأرض، ولا تستخدم للتنقل؛ وبدلا من ذلك فهي تستخدم في الافتراس. سمح اكتشاف الطرف الأمامي الكامل للعينة (NCSM 14345) بإجراء التحليل الأول لوظيفة الطرف الأمامي ومدى حركة في الأكروكانثوصور.[27] فحصت الدراسة أسطح العظام التي قد تكون مفصلية مع عظام أخرى لتحدد مدى حركة المفاصل دون خلع. لم تتطابق العظام معا في كثير من المفاصل بشكل تام، مما يدل على وجود كمية كبيرة من الغضاريف في المفاصل، كما يظهر في العديد من الأركوصورات الحالية. من بين النتائج الأخرى فقد بينت الدراسة أن الأطراف الأمامية كانت تتدلى خلال وضعية الراحة من الكتفين مع انحناء عظم العضد إلى الخلف قليلاً، والكوع منحني وتتجه المخالب باتجاه وسطي (إلى الداخل).[27]

يعتبر كتف الأكروكانثوصور محدودا في الحركة مقارنةً بكتف البشر. ولا يمكن للذراع أن تتأرجح في دائرة كاملة، ولكن يمكن أن يتراجع (التأرجح للخلف) 109 درجة من الزاوية العمودية، بحيث يمكن أن يميل عظم العضد إلى الأعلى قليلاً. وتقتصر الإطالة (التأرجح للأمام) على 24 درجة فقط من الزاوية العمودية. ولا يمكن للذراع الوصول إلى الوضع الرأسي عند التقريب (التأرجح للأسفل) ولكن يمكنه التقريب (التأرجح للأعلى) إلى 9 درجات من الزاوية الأفقية. وتعنبر حركة الكوع محدودة أيضا مقارنة بالبشر، بمدى إجمالي 57 درجة فقط للحركة. لا يمكن أن للذراع أن تمتد (أستقامة) بشكل كامل، ولا يمكن أن تنثني (تنحني) بشكل كبير، مع عدم قدرة عظم العضد على تشكيل الزاوية اليمنى مع الساعد. تنغلق الكعبرة والزند (عظام الساعد) معا بحيث لا توجد إمكانية الكب أو الاستلقاء (الالتواء) كما هو الحال في الساعد البشري.[27]
لا تتناسق عظام الرسغ معا بدقة، مما يشير إلى وجود كمية كبيرة من الغضاريف في الرسغ، وهذا من شأنه أن يزيد من صلابتها. جميع الأصابع قادرة على التمدد المفرط (الانحناء للخلف) حتى أنها تكاد تلامس الرسغ. عند ثنيها يتقارب الإصبع الأوسط مع الإصبع الأول بينما ينحرف الإصبع الثالث للداخل. ويحمل الإصبع الأول أكبر مخلب، والذي يتم ثنيه بشكل دائم بحيث ينحني للخلف باتجاه الجانب السفلي من اليد. وبطريقة مماثلة، قد يكون المخلب الأوسط مثنيا بشكل دائم، بينما المخلب الثالث الأصغر قادرا على الانثناء والتمديد.[27]
بعد تحديد نطاق الحركة في مفاصل الأطراف الأمامية فقد استمرت الدراسة في وضع فرضية حول عادات الافتراس للأكروكانثوصور. لا تستطع الأطراف الأمامية التأرجح إلى الأمام بشكل كبير، حتى أنها غير قادرة حتى على خدش رقبة الحيوان. وبالتالي، من المحتمل أن الأكروكانثوصور لم يكن يستخدم أطرافه الأمامية في القبضة الأولي على الفريسة وربما كان يستخدم فمه عند الصيد. من ناحية أخرى، فإن الأطراف الأمامية كانت قادرة على التراجع نحو الجسم بقوة كبيرة، وبمجرد القبض على الفريسة بالفكين تقوم الأطراف الأمامية القوية بمهمة السحب وإمساك الفريسة بإحكام ومنعها من الهروب. كما أن محاولة الفريسة بالابتعاد مستحيلة بسبب تعلقها بالمخالب المثنية بشكل دائم لأول إصبعين. قد تكون قابلية التمدد المفرطة للأصابع تكيفت لتسمح للأكروكانثوصور بالاحتفاظ بالفريسة تكافح دون الخوف من أن تخلع الأصابع. وبمجرد ضم الفريسة قد يكون الأكروكانثوصور قد اطبق عليها بفكيه. يقول الاحتمال الآخر بأن الأكروكانثوصور قد يحتجز فريسته بفكيه، بينما يسحب أطرافه الأمامية بشكل متكرر لتنتج جروح كبيرة بسبب مخالبه.[27] اقترحت نظريات أخرى لكنها أقل احتمالا بأن يكون نطاق الحركة للأطراف الأمامية قادرا على الإمساك بجانب الفريسة والتشبث بإسقاط الفرائس ذات القامات الأصغر، وبالرغم من أن هذا غير مرجح بسبب امتلاك الأكروكانثوصور لهيكل ساق قوي إلى حد ما مقارنة بوحشيات الأرجل الأخرى ذات البنية المماثلة.
هيكل الدماغ والأذن الداخلية

في 2005، أعاد العلماء بناء قالب داخلي (طبق الأصل) لتجويف جمجمة الأكروكانثوصور باستخدام تصوير مقطعي محوسب (CT scan) لتحليل الفراغات داخل جمجمة العينة النمطية (OMNH 10146). في الطبيعة الحية يكون الكثير من هذه المساحات ممتلئة بالسحايا والسائل الدماغي النخاعي، بالإضافة إلى المخ نفسه. لكن، بالإمكان تحديد السمات العامة للدماغ والأعصاب القحفية من داخل القالب الداخلي ومقارنتها بوحشيات الأرجل الأخرى التي تم إنشاء قوالب داخلية من أجلها. بينما الدماغ مشابه للعديد من وحشيات الأرجل، وأكثرها شبها في الألوصورات. وتشبه إلى حد كبير أدمغة الكاركارودونتوصور والجيجانوتوصور أكثر من أدمغة الألوصور أو السينرابتور، وهذا يعطي الدعم لفرضية أن الأكروكانثوصور كان من الكاركارودونتوصوريدات.[28]
بشبه الدماغ حرف S، دون توسع في نصف الكرة المخية، وهي في التماسيح أكثر من الطيور. هذا يتماشى مع التحفظ الشاملة لأدمغة وحشيات الأرجل الغير سيلوروصورية. لدى الأكروكانثوصور بصيلات شمية كبيرة ومنتفخة، مما يدل على قوة حاسة الشم لديه. يتبين من التركيب المعاد بناءة للقنوات الهلالية في الأذن والتي تتحكم في التوازن أن الرأس مرتبط بزاوية 25 درجة أسفل المستوى الأفقي. تم تحديد ذلك من خلال توجيه القالب الداخلي بحيث تكون القناة الهلالية الجانبية موازية للأرض، كما هو الحال عادة عندما يكون الحيوان في حالة تأهب.[28]
آثار الأقدام المحتملة

يوجد في تكوين غلين روز الواقع في وسط تكساس على العديد من آثار أقدام الديناصورات المحفوظة، بما في ذلك طبعات وحشيات الأرجل الكبيرة ثلاثية الأصابع. تم اكتشاف أشهر هذه المسارات على طول نهر بالوكسى في منتزه ديناصور فالي الحكومي، وقسم منها معروض الآن في المتحف الأمريكي للتاريخ الطبيعي في مدينة نيويورك.[29] ومن المستحيل تحديد نوع الحيوان الذي صنع هذه البصمات لأنه لا توجد عظام أحفورية مرتبطة بالمسارات. ومع ذلك، اعتبر العلماء أنه يحتمل أن آثار الأقدام تنتمي إلى الأكروكانثوصور.[30] تمت دراسة أجريت عام 2001 لمقارنة آثار الأقدام في غلين روز بأقدام العديد من وحشيات الأرجل الكبيرة، لكنها لم تستطع تخصيصها لأي جنس معين. ومع ذلك، فقد أشارت الدراسة إلى أن المسارات كانت ضمن نطاقي الحجم والشكل المتوقع للأكروكانثوصور. لأن تكوين غلين روز قريب من تكويني «أنتلرز» و«جبال التوأم» سواء في الموقع الجغرافي أوالعصر الجيولوجي، ووحشيات الأرجل الوحيدة المعروفة من تلك التكوينات هي الأكروكانثوصورات، وقد خلصت الدراسة إلى أن الأكروكانثوصورات على الأرجح هي من صنع تلك المسارات.[31]
يحتوي مسار غلين روز الشهير المعروض في مدينة نيويورك آثارا لأقدام وحشيات الأرجل تعود إلى عدد منها تحركت في نفس الاتجاه وكذلك اثني عشر ديناصورا من سحليات الأرجل. أحيانًا تكون آثار وحشيات الأرجل فوق آثار أقدام سحليات الأرجل، وتبين أنها قد تشكلت في وقت لاحق. ويقدم هذا دليل على أن مجموعة صغيرة من الأكروكانثوصورات كانت تطارد قطيعا من سحليات الأرجل.[29] وعلى أن هذه الفرضية مثيرة للاهتمام ومعقولة إلا أنه من الصعب إثباتها وتوجد تفسيرات أخرى لذلك. علي سبيل المثال، يحتمل أنه قد تحركت عدة ديناصورات من وحشيات الأرجل بشكل انفرادي في نفس الاتجاه في أوقات مختلفة بعد مرور سحليات الأرجل، مما أدى إلى ظهور قطيع يطارد فريسته. ويمكن قول الشيء نفسه عن «القطيع» المتوقع لسحليات الأرجل، التي ربما كانت أو لم تكن تتحرك كمجموعة.[32] عند تقاطع طريق مسارات لأحد سحليات الأرجل مع أحد المسارات لوحشيات الأرجل تفقد بصمة القدم التي يستشهد بها كدليل على الهجوم.[33] لكن، علماء آخرون يشككون في صحة هذا التفسير لأن سحليات الأرجل لم تغير مشيتها كما هو متوقع إذا كان مفترس كبير معلقًا على جانبها.[32]
اقرأ أيضا
االمراجع
- Stovall، J. Willis؛ Langston, Wann . (1950). "Acrocanthosaurus atokensis, a new genus and species of Lower Cretaceous Theropoda from Oklahoma". American Midland Naturalist. ج. 43 ع. 3: 696–728. DOI:10.2307/2421859. JSTOR:2421859. مؤرشف من الأصل في 2022-03-28.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: أسماء عددية: قائمة المؤلفين (link) صيانة الاستشهاد: أسماء متعددة: قائمة المؤلفين (link) - Bates, K.T.؛ Manning, P.L.؛ Hodgetts, D.؛ Sellers, W.I. (2009). Beckett، Ronald (المحرر). "Estimating Mass Properties of Dinosaurs Using Laser Imaging and 3D Computer Modelling". PLOS ONE. ج. 4 ع. 2: e4532. Bibcode:2009PLoSO...4.4532B. DOI:10.1371/journal.pone.0004532. PMC:2639725. PMID:19225569.
We therefore suggest 5750–7250 kg represents a plausible maximum body mass range for this specimen of Acrocanthosaurus.
- Liddell، Henry George؛ Robert Scott (1980). Greek–English Lexicon, Abridged Edition. Oxford: Oxford University Press. ISBN:978-0-19-910207-5. مؤرشف من الأصل في 2021-08-17.
- Langston، Wann R. (1947). A new genus and species of Cretaceous theropod dinosaur from the Trinity of Atoka County, Oklahoma. Unpublished M.S. thesis. University of Oklahoma.
- Czaplewski، Nicholas J.؛ Cifelli, Richard L.؛ Langston, Wann R. Jr. (1994). "Catalog of type and figured fossil vertebrates. Oklahoma Museum of Natural History". Oklahoma Geological Survey Special Publication. ج. 94 ع. 1: 1–35.
- Harris، Jerald D. (1998). "A reanalysis of Acrocanthosaurus atokensis, its phylogenetic status, and paleobiological implications, based on a new specimen from Texas". New Mexico Museum of Natural History and Science Bulletin. ج. 13: 1–75.
- Currie، Philip J.؛ Carpenter, Kenneth (2000). "A new specimen of Acrocanthosaurus atokensis (Theropoda, Dinosauria) from the Lower Cretaceous Antlers Formation (Lower Cretaceous, Aptian) of Oklahoma, USA". Geodiversitas. ج. 22 ع. 2: 207–246. مؤرشف من الأصل في 2007-11-14.
- D'Emic، Michael D.؛ Melstrom, Keegan M.؛ Eddy, Drew R. (2012). "Paleobiology and geographic range of the large-bodied Cretaceous theropod dinosaur Acrocanthosaurus atokensis". Palaeogeography, Palaeoclimatology, Palaeoecology. 333–334: 13–23. Bibcode:2012PPP...333...13D. DOI:10.1016/j.palaeo.2012.03.003.
- Therrien، F.؛ Henderson, D.M. (2007). "My theropod is bigger than yours...or not: estimating body size from skull length in theropods" (PDF). Journal of Vertebrate Paleontology. ج. 27 ع. 1: 108–115. DOI:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2. مؤرشف من الأصل (PDF) في 2021-06-24.
- Holtz، Thomas R.؛ Molnar, Ralph E.؛ Currie, Philip J. (2004). "Basal Tetanurae". في Weishampel, David B. ؛ بيتر دودسون؛ Osmólska, Halszka (المحررون). The Dinosauria (ط. Second). Berkeley: University of California Press. ص. 71–110. ISBN:978-0-520-24209-8. مؤرشف من الأصل في 2022-02-23.
{{استشهاد بكتاب}}: صيانة الاستشهاد: أسماء عددية: قائمة المحررين (link) صيانة الاستشهاد: أسماء متعددة: قائمة المحررين (link) صيانة الاستشهاد: علامات ترقيم زائدة (link) - Molnar، Ralph E.؛ Kurzanov, Sergei M.؛ Dong Zhiming (1990). "Carnosauria". في Weishampel, David B. ؛ بيتر دودسون؛ Osmólska, Halszka (المحررون). The Dinosauria (ط. First). Berkeley: University of California Press. ص. 169–209. ISBN:978-0-520-06727-1.
{{استشهاد بكتاب}}: صيانة الاستشهاد: أسماء عددية: قائمة المحررين (link) صيانة الاستشهاد: أسماء متعددة: قائمة المحررين (link) صيانة الاستشهاد: علامات ترقيم زائدة (link) - Naish، Darren؛ Hutt, Stephen؛ Martill, David M. (2001). "Saurischian Dinosaurs 2: Theropods". Dinosaurs of the Isle of Wight. London: The Palaeontological Association. ص. 242–309. ISBN:978-0-901702-72-2.
- Brusatte، Stephen L.؛ Benson، Roger B. J.؛ Chure، Daniel J.؛ Xu، Xing؛ Sullivan، Corwin؛ Hone، David W. E. (2009). "The first definitive carcharodontosaurid (Dinosauria: Theropoda) from Asia and the delayed ascent of tyrannosaurids" (PDF). Naturwissenschaften. ج. 96 ع. 9: 1051–8. Bibcode:2009NW.....96.1051B. DOI:10.1007/s00114-009-0565-2. hdl:20.500.11820/33528c2e-0c9c-4160-8693-984f077ee5d0. PMID:19488730. S2CID:25532873. مؤرشف من الأصل (PDF) في 2021-08-27.
- Benson، Roger B. J.؛ Carrano، Matthew T.؛ Brusatte، Stephen L. (2009). "A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic" (PDF). Naturwissenschaften. ج. 97 ع. 1: 71–8. Bibcode:2010NW.....97...71B. DOI:10.1007/s00114-009-0614-x. PMID:19826771. S2CID:22646156. مؤرشف من الأصل (PDF) في 2021-08-27.
- Romer، Alfred S. (1956). Osteology of the Reptiles. Chicago: University of Chicago Press. ص. 772pp. ISBN:978-0-89464-985-1.
- Walker، Alick D. (1964). "Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. ج. 248 ع. 744: 53–134. Bibcode:1964RSPTB.248...53W. DOI:10.1098/rstb.1964.0009.
- Romer، Alfred S. (1966). Vertebrate Paleontology (ط. Third). Chicago: University of Chicago Press. ص. 468pp. ISBN:978-0-7167-1822-2. مؤرشف من الأصل في 2020-07-24.
- Carroll، Robert L. (1988). Vertebrate Paleontology and Evolution. W.H. Freeman and Company. ISBN:978-0-7167-1822-2. مؤرشف من الأصل في 2021-07-14.
- Lambert، David؛ Diagram Group (1983). "Spinosaurids". A Field Guide to Dinosaurs. New York: Avon Books. ص. 84–85. ISBN:978-0-380-83519-5.
- Norman، David B. (1985). "Carnosaurs". The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom. New York: Crescent Books. ص. 62–67. ISBN:978-0-517-46890-6.
- Glut، Donald F. (1982). The New Dinosaur Dictionary. Secaucus, NJ: Citadel Press. ص. 39, 48. ISBN:978-0-8065-0782-8. مؤرشف من الأصل في 2020-07-24.
- Paul، Gregory S. (1988). "Genus Acrocanthosaurus". Predatory Dinosaurs of the World. New York: Simon & Schuster. ص. 314–315. ISBN:978-0-671-61946-6.
- Olshevsky، George (1991). A Revision of the Parainfraclass Archosauria Cope, 1869, Excluding the Advanced Crocodylia. San Diego: Publications Requiring Research. ص. 196pp.
- Eddy، Drew R.؛ Clarke، Julia A. (2011). Farke، Andrew (المحرر). "New Information on the Cranial Anatomy of Acrocanthosaurus atokensis and Its Implications for the Phylogeny of Allosauroidea (Dinosauria: Theropoda)". بلوس ون. ج. 6 ع. 3: e17932. Bibcode:2011PLoSO...617932E. DOI:10.1371/journal.pone.0017932. PMC:3061882. PMID:21445312.
- Novas، Fernando E. (2013). "Evolution of the carnivorous dinosaurs during the Cretaceous: The evidence from Patagonia". Cretaceous Research. ج. 45: 174–215. DOI:10.1016/j.cretres.2013.04.001.
- D'Emic، Michael؛ Melstrom، Keegan؛ Eddy، Drew (15 مايو 2012). "Paleobiology and geographic range of the large-bodied Cretaceous theropod dinosaur Acrocanthosaurus atokensis". Palaeogeography, Palaeoclimatology, Palaeoecology. 333–334: 13–23. Bibcode:2012PPP...333...13D. DOI:10.1016/j.palaeo.2012.03.003.
- Senter، Phil؛ Robins, James H. (2005). "Range of motion in the forelimb of the theropod dinosaur Acrocanthosaurus atokensis, and implications for predatory behaviour". Journal of Zoology. ج. 266 ع. 3: 307–318. DOI:10.1017/S0952836905006989.
- Franzosa، Jonathan؛ Rowe, Timothy. (2005). "Cranial endocast of the Cretaceous theropod dinosaur Acrocanthosaurus atokensis". Journal of Vertebrate Paleontology. ج. 25 ع. 4: 859–864. DOI:10.1671/0272-4634(2005)025[0859:CEOTCT]2.0.CO;2. مؤرشف من الأصل في 2016-02-24.
- Bird، Roland T. (1941). "A dinosaur walks into the museum". Natural History. ج. 43: 254–261.
- Langston، Wann (1974). "Non-mammalian Comanchean tetrapods". Geoscience and Man. ج. 3: 77–102.
- Farlow، James O. (2001). "Acrocanthosaurus and the maker of Comanchean large-theropod footprints". في Tanke, Darren ؛ كينيث كاربنتر (المحررون). Mesozoic Vertebrate Life. Bloomington: Indiana University Press. ص. 408–427. ISBN:978-0-253-33907-2.
{{استشهاد بكتاب}}: صيانة الاستشهاد: أسماء عددية: قائمة المحررين (link) صيانة الاستشهاد: أسماء متعددة: قائمة المحررين (link) صيانة الاستشهاد: علامات ترقيم زائدة (link) - Lockley، Martin G. (1991). Tracking Dinosaurs: A New Look at an Ancient World. Cambridge: Cambridge University Press. ص. 252pp. ISBN:978-0-521-39463-5.
- Thomas، David A.؛ Farlow, James O. (1997). "Tracking a dinosaur attack". Scientific American. ج. 266 ع. 6: 48–53. Bibcode:1997SciAm.277f..74T. DOI:10.1038/scientificamerican1297-74.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
.jpg.webp)













 بوابة الولايات المتحدة
بوابة الولايات المتحدة بوابة ديناصورات
بوابة ديناصورات بوابة علم الأحياء التطوري
بوابة علم الأحياء التطوري بوابة علم الأحياء القديمة
بوابة علم الأحياء القديمة بوابة علم الحيوان
بوابة علم الحيوان