Sauropoda
Sauropoda, of die souropodes ("met akkedisvoete"), was ’n infraorde van Saurischia-dinosourusse. Hulle het baie lang nekke en sterte, klein koppe (in vergelyking met die res van hul lyf) en dik, bonkige bene gehad. Hulle is merkwaardig vanweë die enorme grootte van sommige spesies – die groep sluit die grootste diere in wat op aarde geloop het. Bekende genera sluit in Brachiosaurus, Diplodocus en Apatosaurus (wat volgens die huidige klassifikasie ’n vroeëre sinoniem vir Brontosaurus) is. Souropodes het in die Laat Trias-periode vir die eerste keer hul verskyning gemaak. Teen die Laat Jura (150 miljoen jaar gelede) het souropodes wydverspreid voorgekom en teen die Laat Kryt is hulle hoofsaaklik vervang deur titanosourusse, wat feitlik wêreldwyd voorgekom het. Laasgenoemde het saam met alle ander nie-vlieënde dinosourusse uitgesterf tydens die Kryt-Paleogeen-uitwissing. Fossiele oorblyfsels van souropodes is op al die kontinente ontdek, insluitende Antarktika.
| Sauropoda | |
|---|---|
 | |
| Die skelet van ’n Apatosaurus louisae (Carnegie-museum van Natuurgeskiedenis). | |
| Wetenskaplike klassifikasie | |
| Koninkryk: | |
| Filum: | |
| Subfilum: | |
| Klas: | |
| Superorde: | |
| Orde: | |
| Suborde: | |
| Infraorde: | †Sauropoda (Marsh, 1878) |
| Sauropoda | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Kladistiese klassifikasie | ||||||||||||
| ||||||||||||
| Subgroepe | ||||||||||||
| ||||||||||||
Die naam is in 1878 deur die Amerikaanse paleontoloog O.C. Marsh gevestig en is afgelei van die Grieks vir "akkedisvoet".[1] Souropodes is een van die mees herkenbare groepe dinosourusse en word algemeen voorgestel in verskeie kunsvorms vanweë hul grootte.
Volledige souropode-fossiele is skaars. Baie spesies, veral die grootstes, is net bekend uit geïsoleerde stukke bene. Baie feitlik volledige fossiele kom koppe, sterte of ledemate kort.
Beskrywing
Souropodes was plantvretende, vierpotige diere met meestal lang nekke,[2] dikwels met roerspaanvormige tande (breed by die basis en smaller bo). Hulle het klein koppe en massiewe lywe gehad, en die meeste se sterte was lank. Hul agterbene was dik, reguit en kragtig. Hulle het vyf tone gehad, waarvan die binneste drie (of soms vier) kloue gehad het. Hul voorste ledemate was dunner en het in pilaaragtige pote geëindig wat hul gewig kon steun; net die duim het ’n klou gehad.
Grootte

Amphicoelias fragillimus Argentinosaurus huinculensis Mamenchisaurus sinocanadorum Supersaurus vivianae Sauroposeidon proteles
Die souropodes se mees bekende eienskap was hul grootte. Selfs die dwerg-souropodes soos Europasaurus (moontlik 5 tot 6 meter lank) was van die grootste diere in hul onderskeie ekosisteme. Hul enigste mededingers wat grootte betref, was vinwalvisse soos die blouwalvis. Maar anders as walvisse het hulle hoofsaaklik op land voorgekom.
Hul liggaamstruktuur het nie so baie gewissel soos dié van ander dinosourusse nie, moontlik vanweë groottebeperkings, maar daar was tog variasies. Sommige, soos die Diplodocidae, het uiters lang sterte gehad, wat hulle moontlik soos ’n sweep kon klap om hul aanvallers af te skrik of te beseer.[3] Supersaurus, wat 33 tot 34 m lank was,[4] was die langste souropode waarvan ’n feitlik volledige skelet ontdek is, maar die ou rekordhouer, Diplodocus, was ook baie lank. Die holotipe-ruggraat van Amphicoelias fragillimus, wat nou weg is, kon dié gewees het van ’n dier van 58 m lank;[5] sy werwelkolom sou veel langer gewees het as dié van die blouwalvis. Die langste moderne dier op land, die luislang Python reticulatus, word net sowat 8,7 m lank.[6]
Ander, soos die Brachiosauridae, was uiters hoog, met hoë skouers en uiters lang nekke. Sauroposeidon was moontlik die hoogste – tot sowat 18 m. In vergelyking daarmee word die kameelperd, die hoogste van alle lewende diere, net 4,8 tot 5,5 m hoog.
Sommige souropodes was ongelooflik swaar – Argentinosaurus was moontlik die swaartse: 80 tot 100 ton. Paralititan, Antarctosaurus en Argyrosaurus was amper net so swaar. Daar is skamele getuienis van ’n moontlik swaarder titanosouriër, Bruhathkayosaurus, wat tussen 175 en 220 ton kon geweeg het. Die grootste lewende landdier, die savanne-olifant, weeg nie meer as 10 ton nie.
Onder die kleinste souropodes tel die primitiewe Ohmdenosaurus (4 m lank), die dwerg-titanosourus Magyarosaurus (5,3 m) en die dwerg-brachiosouride Europasaurus, wat as ’n volgroeide dier 6,2 m lank was.[7] Ook noemenswaardig is die Brachytrachelopan, wat die kortste lid van die groep was vanweë sy besonder kort nek. Anders as ander souropodes, waarvan die nek tot vier keer so lank as die rug was, was die Brachytrachelopan se nek korter as sy rug.
Ledemate
As enorme, swaar viervoetiges het souropodes gespesialiseerde ledemate ontwikkel wat hul gewig sou kon dra. Die agterbene was breed en die meeste spesies het drie kloue agter gehad.[8] Wat veral van ander diere verskil het, was die hoogs aangepaste voorpote (manus). Dit het glad nie gelyk soos dié van moderne vierpotiges, soos die olifant, nie. In plaas van uitsprei na die kante, was souropodes se manus-bene gerangskik in ten volle vertikale kolomme, met uiters kort vingerbene.[9]
Die rangskikking van die voorpootkolomme was halfsirkelvormig en daarom het souropodes se voetafdrukke soos hoefysters gelyk. Hulle het waarskynlik nie vlesige kussings gehad soos die olifant nie en die voorpote was dus konkaaf.[9] Die enigste klou wat sigbaar was, was die besonderse duimklou. Feitlik alle souropodes het hulle gehad, hoewel die doel daarvan onbekend is.
Latere titanosourusse het nie die duimklou gehad nie; dié diere was ongewoon vir souropodes aangesien hulle ook geen tone aan die voorpote gehad het nie. Hulle het op "stompies" geloop.[10]
Lugsakke
Nes ander Saurischia-dinosourusse (soos voëls en ander teropodes), het souropodes ’n stelsel lugsakke gehad – dit is duidelik uit holtes in die meeste van hul werwels.[11]
Die voëlagtige holtes in souropodes se bene is vroeg reeds ontdek; minstens een souropode-fossiel wat in die 19de eeu ontdek is, Ornithopsis, is as gevolg daarvan aanvanklik verkeerd geklassifiseer as ’n pterosourus (’n vlieënde reptiel).[12]
Pantsers

Sommige souropodes het pantsers gehad. Daar was genera met klein knotse aan hul sterte, soos Shunosaurus, en verskeie titanosourusse soos Saltasaurus en Ampelosaurus het klein, skubberige osteoderms gehad wat dele van hul lywe bedek het.
Tande
In ’n studie deur Michael D’Emic en sy kollegas van Stony Brook-universiteit is bevind souropodes het dikwels tande gewissel vanweë hul groot aptyt. Volgens die studie is elke tand van Nigersaurus byvoorbeeld elke 14 dae vervang, dié van Diplodocus elke 35 dae en dié van Camarasaurus elke 62 dae.[13] Die wetenskaplikes het bevind sekere eienskappe van die tande bepaal hoe lank hulle vat om terug te groei; Camarasaurus se tande het langer gevat as dié van Diplodocus omdat hulle groter was.[14]
D'Emic en sy span het ook bevind die verskille in tande dui op verskillende diëte. Diplodocus het plante naby die grond gevreet, terwyl Camarasaurus blare van die boonste en middelste boomtakke gevreet het. Volgens die wetenskaplikes het die gespesialiseerde diëte die verskillende plantvretende dinosourusse in staat gestel om saam te bestaan.[13][14]
Paleobiologie
Ekologie
Toe souropodes vir die eerste keer ontdek is, het wetenskaplikes hulle met moderne walvisse vergelyk vanweë hul enorme grootte. In die meeste studies van die 19de en vroeë 20ste eeu is bevind souropodes was te swaar om hul eie gewig op land te kon dra en daarom moes hulle waterdiere gewees het. Die meeste kunsvoorstellings van die diere regdeur die eerste driekwart van die 20ste eeu is van waar die diere heeltemal of gedeeltelik met water bedek is.[15] Aan die begin van die 1950's het wetenskaplikes begin twyfel aan dié teorie toe ’n studie deur Kermack (1951) gewys het die druk van die water sou die longe en lugweg inmekaargedruk het as die dier ’n paar meter diep in die water was.[16] In dié studie is bewyse dat souropodes talle lugsakke gehad het, egter geïgnoreer.
In die vroeë 1970's is die uitwerking van souropodes se lugsakke op hul veronderstelde seelewe begin ondersoek. Paleontoloë soos Coombs en Bakker het dit gebruik om te bewys souropodes was hoofsaaklik landdiere. In 2004 het D.M. Henderson daarop gewys dat souropodes vanweë hul lugsakke nie heeltemal in die water sou kon afsak nie en dat hulle dus bo-op sou dryf; daarom sou die water nie hul longe inmekaardruk nie.[15]
Vanweë hul liggaamsproporsies sou drywende souropodes egter baie onstabiel gewees het en hulle was nie aangepas om vir lang tye in die water te bly nie.[15] Al word daar nie meer geglo souropodes was waterdiere nie, is daar getuienis dat hulle wel nat omgewings en kushabitats verkies het. Voetspore is dikwels aan kuslyne of oor vloedvlaktes gevind, en souropode-fossiele word dikwels ontdek in nat omgewings of saam met fossiele van see-organismes.[15]
Tropvorming en ouerskap
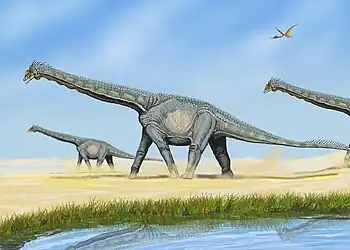
Daar is baie getuienis dat souropodes kuddediere was wat troppe gevorm het. Die soort troppe het egter van spesie tot spesie verskil. Op sommige terreine, soos in Argentinië, is bewyse uit die Middel-Jura gevind dat individue van verskillende ouderdomsgroepe – volgroeide en jong diere – saam troppe gevorm het. Op ander terreine is voorbeelde gevind van souropodes wat in troppe gereis het wat volgens ouderdom geskei was, met jong en volgroeide diere in apart groepe. Sulke geskeide troppe is gevind met betrekking tot spesies soos Alamosaurus, Bellusaurus en sommige lede van die familie Diplodocidae.[17]
Myers en Fiorillo het in ’n studie bevind volgroeide en jong souropodes se tande verskil en dit dui op verskillende diëte, en daarom sou dit doeltreffender gewees het om in aparte groepe te reis. Die enorme verskille in grootte sou ook ’n rol kon gespeel het.[17]
Aangesien die skeiding moes plaasgevind het kort nadat die kleintjies uitgebroei het, tesame met bewyse dat souropodes vroegselfstandig was, het Myers en Fiorillo tot die gevolgtrekking gekom dat spesies met geskeide troppe nie veel tekens van ouersorg getoon het nie.[17] Wetenskaplikes wat gemengde groepe bestudeer het, het weer tot die gevolgtrekking gekom dat sulke spesies vir ’n lang tyd na hul kleintjies omgesien het.[18]
Dit is onbekend in hoe ’n mate die voorkoms van gemengde en geskeide troppe van groep tot groep gewissel het. Nog voorbeelde van kuddevorming moet ontdek word om patrone te kan onderskei.[17]
Regop houding

Wetenskaplikes soos Henry Fairfield Osborn het vroeg reeds vermoed souropodes kon op hul agterpote staan en hul stert gebruik as die derde "poot" van ’n driepoot.[19] In ’n 2005-verslag het Rothschild en Molnar geredeneer as souropodes soms op hul agterpote kon staan, sou daar bewyse moes wees van stresfrakture in die voorpote. Geen sulke bewyse is egter gevind nie nadat hulle ’n groot verskeidenheid souropode-skelette ondersoek het.[20]
Heinrich Mallison was in 2009 die eerste wetenskaplike wat die fisieke potensiaal van verskeie souropodes ondersoek het om op hul agterpote te kan staan. Hy het bevind sekere eienskappe wat voorheen verbind is met so ’n houding, het eintlik geen verband daarmee gehad nie (soos die breë heupbene van titanosourusse) of sou dit eintlik belemmer het. Titanosourusse het byvoorbeeld ’n baie buigbare ruggraat gehad wat hom onstabiel sou maak as hy op sy agterpote staan en meer stremming op die spiere sou plaas.
Dit is ook onwaarskynlik dat brachiosourides op hul agterpote kon staan, aangesien hul swaartekragmiddelpunt veel verder vorentoe was as dié van ander souropodes en dit sou hulle in so ’n houding onstabiel gemaak het.[21]
Diplodosides was aan die ander kant blykbaar beter aangepas vir ’n driepoothouding. Hul swaartekragmiddelpunt was reg bo die heupe en dit sou hulle genoeg balans gegee het om op twee pote te staan. Hulle het ook die mees beweeglike nekke van alle souropodes gehad, asook ’n goed gespierde heupgordel en stertwerwels met ’n gespesialiseerde vorm sodat die stert gewig sou kon dra op die punt waar dit aan die grond geraak het.
Mallison het gereken diplodosides was beter aangepas om regop te staan as olifante, wat dit soms in die natuur doen.[21]
Grootte-evolusie
Verskeie wetenskaplikes het probeer uitvind hoekom souropodes so groot geword het. Dit het al vroeg in die diere se evolusie gebeur, selfs met die verskyning van die eerste ware souropodes in die Laat Trias-periode. Volgens Kenneth Carpenter moes die faktor wat tot die grootte van dié diere gelei het, van die begin van die groep se ontstaan reeds teenwoordig gewees het.[5]
Studies van plantvretende soogdiere wat baie groot word, soos olifante, toon groter diere verteer kos beter. Omdat hul spysverteringstelsel langer is, bly kos aansienlik langer daarin en daarom kan diere op kos van laer gehalte oorleef. Gedurende souropodes se hele evolusiegeskiedenis is hulle hoofsaaklik in halfdroë gebiede aangetref wat in sekere seisoene droër was, en dit het tot ’n laer kosgehalte in dié seisoene gelei. Die omgewing van die meeste souropodes van die Laat Jura was hoofsaaklik savanne, en dit ondersteun die teorie dat swak kosgehalte lei tot die evolusie van reusagtige plantvreters.[5]
Ontdekkingsgeskiedenis
Die eerste stukke fossiele oorblyfsels wat nou as dié van souropodes erken word, kom almal uit Engeland en is aanvanklik op verskeie manier vertolk. Hul verwantskap met ander dinosourusse sou eers lank ná hul ontdekking besef word.


Die eerste souropode-fossiel wat wetenskaplik beskryf is, was ’n enkele tand wat die naam Rutellum implicatum gekry het.[22] Die fossiel is in 1699 deur die Wallieser Edward Lhuyd beskryf, maar is nie destyds herken as ’n reusagtige prehistoriese reptiel nie.[23] Dinosourusse sou eers meer as ’n eeu later as ’n groep erken word.
Die Engelse paleontoloog Richard Owen het in 1841 die eerste moderne wetenskaplike beskrywing van souropodes gepubliseer in sy verslag oor Cetiosaurus en Cardiodon. Cardiodon was net bekend uit twee ongewone, hartvormige tande (vandaar sy naam). Al wat van die tande bekend was, was dat hulle behoort het aan ’n voorheen onbekende, groot reptiel. Cetiosaurus was bekend uit effens beter, maar steeds brokkige oorblyfsels. Owen het destyds gedink Cetiosaurus was ’n reusagtige seereptiel wat aan die moderne krokodil verwant was, vandaar sy naam ("walvisakkedis"). ’n Jaar later, toe Owen die naam Dinosauria gevestig het, het hy nie Cetiosaurus en Cardiodon in dié groep ingesluit nie.[24]
In 1850 het Gideon Mantell die verwantskap besef tussen dinosourusse en verskeie bene wat Owen as Cetiosaurus geklassifiseer het. Hy het opgelet die bene in die dier se pote het ’n murgholte, wat kenmerkend is van landdiere. Hy het hulle onder die nuwe genus Pelorosaurus geklassifiseer saam met dinosourusse. Mantell het egter steeds nie die verwantskap met Cetiosaurus besef nie.[12]
Die volgende souropode wat ontdek en verkeerd geklassifiseer is, was ’n stel heupwerwels wat Harry Seeley in 1870 beskryf het. Seeley het gesien die werwels is baie lig vir hul grootte en bevat holtes vir lugsakke. Die enigste bekende diere destyds met lugsakke was voëls en pterosourusse, en Seeley het gemeen die werwels behoort aan ’n pterosourus. Hy het die nuwe genus Ornithopsis ("voëlgesig") genoem.[12]
Toe vollediger oorblyfsels van Cetiosaurus in 1871 deur Phillips beskryf is, het hy eindelik besef die dier is ’n dinosourus wat aan Pelorosaurus verwant is.[25] Dit was egter eers met die ontdekking van byna volledige souropode-skelette (Apatosaurus en Camarasaurus) in die VSA later dié jaar dat die volledige prentjie van souropodes duidelik geraak het. John A. Ryder het ’n benaderde rekonstruksie van ’n volledige souropode-skelet gemaak wat gebaseer was op die oorblyfsels van Camarasaurus, hoewel baie eienskappe steeds onakkuraat of onvolledig was volgens latere vondste en biomeganiese studies.[26] In 1877 het Richard Lydekker nog ’n verwant van Cetiosaurus genoem, naamlik Titanosaurus, gebaseer op ’n enkele werwel.[12]
In 1878 is die volledigste souropode nog gevind. Dit is beskryf deur Othniel Charles Marsh, wat dit Diplodocus genoem het. Met hierdie vonds het Marsh ’n nuwe groep geskep wat Diplodocus, Cetiosaurus en hul toenemende aantal verwante bevat om hulle te onderskei van ander groot groepe dinosourusse. Marsh het die groep Sauropoda ("akkedisvoete") genoem.[12]
Klassifikasie
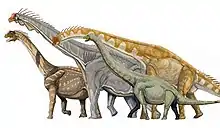
Die wetenskaplike klassifikasie van die souropodes is redelik volledig, hoewel daar nog onsekerhede is, soos die plek van Euhelopus, Haplocanthosaurus, Jobaria en Nemegtosauridae.
Die volgende kladogram is geskoei op ’n ontleding deur Apaldetti en sy kollegas in 2011.[27]
| Sauropoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Verwysings
- Marsh, O.C. (1878). "Principal characters of American Jurassic dinosaurs. Part I"". American Journal of Science and Arts. 16: 411–416.
- Michael P. Taylor, Mathew J. Wede (2013). "Why sauropods had long necks; and why giraffes have short necks". PeerJ. 1: e36. doi:10.7717/peerj.36. PMC 3628838. PMID 23638372.
{{cite journal}}: AS1-onderhoud: gebruik authors-parameter (link) - Bakker, Robert (1994). "The Bite of the Bronto" Earth 3:(6):26–33.
- Lovelace, David M.; Hartman, Scott A.; Wahl, William R. (2007). "Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny". Arquivos do Museu Nacional. 65 (4): 527–544.
- Carpenter, K. (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus." In Foster, J.R. and Lucas, S.G., eds., 2006, Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 131–138.
- Murphy JC, Henderson RW. 1997. Tales of Giant Snakes: A Historical Natural History of Anacondas and Pythons. Krieger Pub. Co. 221 pp. ISBN 0-89464-995-7.
- Martin Sander, P.; Mateus, Octávio; Laven, Thomas; Knötschke, Nils (2006). "Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur". Nature. 441 (7094): 739–41. doi:10.1038/nature04633. PMID 16760975.
- Bonnan, M.F. 2005. Pes anatomy in sauropod dinosaurs: implications for functional morphology, evolution, and phylogeny; pp. 346-380 in K. Carpenter and V. Tidwell (reds.), Thunder-Lizards: The Sauropodomorph Dinosaurs. Bloomington, IN: Indiana University Press.
- Bonnan, Matthew F. (2003). "The evolution of manus shape in sauropod dinosaurs: Implications for functional morphology, forelimb orientation, and phylogeny". Journal of Vertebrate Paleontology. 23 (3): 595. doi:10.1671/A1108.
- Apesteguía, S. (2005). "Evolution of the titanosaur metacarpus". Pp. 321-345 in Tidwell, V. and Carpenter, K. (eds.) Thunder-Lizards: The Sauropodomorph Dinosaurs. Indianapolis: Indiana University Press.
- Wedel, M.J. (2009). "Evidence for bird-like air sacs in Saurischian dinosaurs Geargiveer 15 November 2015 op Wayback Machine". (pdf) Journal of Experimental Zoology, 311A: 18pp.
- Taylor, M.P. (In press). "Sauropod dinosaur research: a historical review". In Richard Moody, Eric Buffetaut, David M. Martill and Darren Naish (eds.), Dinosaurs (and other extinct saurians): a historical perspective. HTML abstract.
- D’Emic, Michael D.,Whitlock, John A., Smith, Kathlyn M., Fisher, Daniel C., Wilson, Jeffrey A. (17 Julie 2013). "Evolution of High Tooth Replacement Rates in Sauropod Dinosaurs". PLOS ONE. 8 (7): e69235. doi:10.1371/journal.pone.0069235. PMC 3714237. PMID 23874921.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - Barber, Elizabeth (9 Junie 2004). "No toothbrush required: Dinosaurs replaced their smile every month" (in Engels). Christian Science Monitor. Geargiveer vanaf die oorspronklike op 3 Maart 2020. Besoek op 14 Augustus 2013.
- Henderson, D.M. (2004). "Tipsy punters: sauropod dinosaur pneumaticity, buoyancy and aquatic habits." Proceedings of the Royal Society of London B, 71: S180–S183. doi:10.1098/rsbl.2003.0136
- Kermack, K.A. (1951). "A note on the habits of sauropods". Ann. Mag. Nat. Hist. 4: 830–832.
- Myers, T.S.; Fiorillo, A.R. (2009). "Evidence for gregarious behavior and age segregation in sauropod dinosaurs". Palaeogeography, Palaeoclimatology, Palaeoecology. 274: 96–104.
- Coria, R.A. (1994). "On a monospecific assemblage of sauropod dinosaurs from Patagonia: implications for gregarious behavior". GAIA. 10: 209–213.
- Osborn, H. F. (1899). "A Skeleton of Diplodocus, Recently Mounted in the American Museum". Science. 10 (259): 870–4. doi:10.1126/science.10.259.870. PMID 17788971.
- Rothschild, B.M. and Molnar, R.E. (2005). "Sauropod Stress Fractures as Clues to Activity". In Carpenter, K. and Tidswell, V. (red.). Thunder Lizards: The Sauropodomorph Dinosaurs. Indiana University Press. pp. 381–391. ISBN 0-253-34542-1.
{{cite book}}: AS1-onderhoud: meer as een naam (link) - Mallison, H. (2009). "Rearing for food? Kinetic/dynamic modeling of bipedal/tripodal poses in sauropod dinosaurs". P. 63 in Godefroit, P. en Lambert, O. (reds), Tribute to Charles Darwin and Bernissart Iguanodons: New Perspectives on Vertebrate Evolution and Early Cretaceous Ecosystems. Brussels.
- Delair, J.B.; Sarjeant, W.A.S. (2002). "The earliest discoveries of dinosaurs: the records re-examined". Proceedings of the Geologists' Association. 113: 185–197. doi:10.1016/S0016-7878(02)80022-0.
- Lhuyd, E. (1699). Lithophylacii Britannici Ichnographia, sive lapidium aliorumque fossilium Britannicorum singulari figura insignium. Gleditsch and Weidmann: London.
- Owen, R. (1842). "Report on British Fossil Reptiles". Part II. Report of the British Association for the Advancement of Science, Plymouth, England.
- Phillips, J. (1871). Geology of Oxford and the Valley of the Thames. Oxford: Clarendon Press, 523 pp.
- Osborn, H.F., and Mook, C.C. (1921). "Camarasaurus, Amphicoelias and other sauropods of Cope". Memoirs of the American Museum of Natural History, n.s. 3:247-387 en plate LX-LXXXV.
- Cecilia Apaldetti, Ricardo N. Martinez, Oscar A. Alcober and Diego Pol (2011). "A New Basal Sauropodomorph (Dinosauria: Saurischia) from Quebrada del Barro Formation (Marayes-El Carrizal Basin), Northwestern Argentina". PLoS ONE. 6 (11): e26964. doi:10.1371/journal.pone.0026964. PMC 3212523. PMID 22096511.
{{cite journal}}: AS1-onderhoud: gebruik authors-parameter (link)
Bronne
- Bob Strauss, 2008, Sauropods: The Biggest Dinosaurs that Ever Lived, The New York Times
- Kristina Curry Rogers en Jeffrey A. Wilson, 2005, The Sauropods: Evolution and Paleobiology, University of California Press, Berkeley, ISBN 0-520-24623-3
- Upchurch, P., Barrett, P.M. en Dodson, P. 2004. Sauropoda. In The Dinosauria, 2de uitg. D. Weishampel, P. Dodson enH. Osmólska (reds.). University of California Press, Berkeley. Pp. 259–322.
Eksterne skakels
 Wikimedia Commons het meer media in die kategorie Sauropoda.
Wikimedia Commons het meer media in die kategorie Sauropoda. Wikispecies het meer inligting verwant aan Sauropoda
Wikispecies het meer inligting verwant aan Sauropoda