Adenosientrifosfaat
Adenosientrifosfaat (afgekort ATP, van die Engelse naam adenosine triphosphate) is 'n organiese verbinding en hidrotroop[Nota 1] wat energie verskaf om baie prosesse in lewende selle aan te dryf, soos spiersametrekking, senuwee-impulsvoortplanting, en chemiese sintese. Dit word in alle bekende lewensvorme gevind en word dikwels na verwys as die "molekulêre geldeenheid" van intrasellulêre energie-oordrag.[2] Wanneer dit in metaboliese prosesse verbruik word, word dit óf na adenosiendifosfaat (ADP) óf na adenosienmonofosfaat (AMP) omgeskakel. Ander prosesse regenereer ATP sodat die menslike liggaam elke dag sy eie liggaamsgewig-ekwivalent in ATP herwin.[3] Dit is ook 'n voorloper van DNS en RNS, en word as 'n koënsiem gebruik.
|
Algemeen | |
|---|---|
| Naam | Adenosientrifosfaat |
| IUSTC-naam | O1-{[(2R,3S,4R,5R)-5-(6-Amieno-9H-purien-9-iel)-3,4-dihidroksioksolaan-2-iel]metiel}tetrawaterstoftrifosfaat |
| Sistematiese naam | Adenosien-5′-(tetrawaterstoftrifosfaat) |
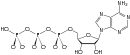 Struktuurformule van Adenosientrifosfaat |
 |
| Chemiese formule | C10H16N5O13P3 |
| Molêre massa | 507,18 g•mol-1 |
| CAS-nommer | 56-65-5 |
| Voorkoms | Kleurlose soute[1] |
| Fasegedrag | |
| Smeltpunt | |
| Kookpunt | |
| Digtheid | 1,04 g•cm-3 |
| Oplosbaarheid | |
|
Suur-basis eienskappe | |
| pKa | 6.5 |
|
Veiligheid | |
| Flitspunt | |
|
Tensy anders vermeld is alle data vir standaardtemperatuur en -druk toestande. | |
| Portaal | |
Uit die perspektief van biochemie word ATP as 'n nukleosiedtrifosfaat[Nota 2] geklassifiseer, wat aandui dat dit uit drie komponente bestaan: 'n stikstofbasis (adenien), die riboos-suiker en die trifosfaat.
Struktuur

ATP bestaan uit 'n adenien wat deur die 9-stikstofatoom aan die 1' koolstofatoom van 'n suiker (ribose) geheg is, wat op sy beurt by die 5' koolstofatoom van die suiker aan 'n trifosfaatgroep geheg is. In sy vele reaksies wat met metabolisme verband hou, bly die adenien- en suikergroepe onveranderd, maar die trifosfaat word omgeskakel na di- en monofosfaat, wat onderskeidelik die afgeleides ADP en AMP gee. Daar word na die drie fosforielgroepe verwys as die alfa (α), beta (β), en, vir die terminale fosfaat, gamma (γ).
In neutrale oplossing bestaan geïoniseerde ATP meestal as ATP4−, met 'n klein deel van ATP3−.[4]
Chemiese eienskappe
ATP is stabiel in waterige oplossings tussen pH 6,8 en 7,4 in die afwesigheid van katalisators. By meer ekstreme pH's hidroliseer dit vinnig na ADP en fosfaat:
- ATP + H2O → ADP + Pi ΔG° = −30,5 kJ/mol
Lewende selle handhaaf die verhouding van ATP tot ADP op 'n punt tien ordes van grootte vanaf ewewig, met ATP-konsentrasies vyf keer hoër as die konsentrasie van ADP.[5][6] In die konteks van biochemiese reaksies word daar gereeld na die P-O-P-bindings verwys as hoë-energiefosfaatbindings.[7]
Soute van ATP kan as kleurlose vaste stowwe geïsoleer word.[1]
Produksie van uit AMP en ADP
Produksie, aërobiese toestande
'n Tipiese intrasellulêre konsentrasie van ATP is moeilik om vas te stel, maar verslae het getoon dat daar 1-10 μmol per gram weefsel in 'n verskeidenheid eukariote is.[8] Die defosforilering van ATP en herfosforilering van ADP en AMP vind herhaaldelik plaas in die verloop van aërobiese metabolisme.
ATP kan deur 'n aantal afsonderlike sellulêre prosesse geproduseer word; die drie metodes in eukariote is (1) glikolise, (2) die sitroensuursiklus/oksidatiewe fosforilering, en (3) beta-oksidasie. Die algehele proses van oksidasie van glukose na koolstofdioksied, die kombinasie van metodes 1 en 2, bekend as sellulêre respirasie, produseer ongeveer 30 ekwivalente van ATP uit elke molekule van glukose.[9]
ATP-produksie deur 'n nie-fotosintetiese aërobiese eukariote vind hoofsaaklik in die mitochondria plaas, wat byna 25% van die volume van 'n tipiese sel uitmaak.[10]
1. Glikolise
In glikolise word glukose en gliserol gemetaboliseer na piruvaat. Glikolise genereer twee ekwivalente van ATP deur substraatfosforilering wat deur twee ensieme, fosfogliseraatkinase en piruvaatkinase, gekataliseer word. Twee ekwivalente van die koënsiem nikotinamiedadeniendinukleotied (NADH) word ook geproduseer, wat via die elektronvervoerketting geoksideer kan word en lei tot die generering van bykomende ATP deur ATP-sintase. Die piruvaat wat as 'n eindproduk van glikolise gegenereer word, is 'n substraat vir die Krebs-siklus.[Nota 3][11]
2. Sitroensuur siklus
In die mitochondria word piruvaat deur die piruvaatdehidrogenase-kompleks geoksideer na die asetielgroep, wat deur die sitroensuursiklus (ook bekend as die Krebs-siklus) volledig tot koolstofdioksied geoksideer word. Elke "draai" van die sitroensuursiklus produseer twee molekules koolstofdioksied, een ekwivalent van ATP-guanosientrifosfaat (GTP) deur substraatvlak-fosforilering wat deur suksinielkoënsiem-A-sintetase gekataliseer word. Suksinielkoënsiem-A word na suksinaat, drie ekwivalente van nikotinamiedadeniendinukleotied (NADH), en een ekwivalent van flavienadeniendinukleotied (FADH2) omgeskakel. NADH en FADH2 word herwin wat bykomende ATP genereer deur oksidatiewe fosforilering. Die oksidasie van NADH lei tot die sintese van 2–3 ekwivalente ATP, en die oksidasie van een FADH2 lewer tussen 1–2 ekwivalente van ATP.[9] Die meerderheid sellulêre ATP word deur hierdie proses gegenereer. Alhoewel die sitroensuursiklus self nie molekulêre suurstof behels nie, is dit 'n verpligte aërobiese proses omdat O2 gebruik word om die NADH en FADH2 te herwin en die chemiese energie verskaf wat die proses dryf.[12] In die afwesigheid van suurstof hou die sitroensuursiklus op.[10]
3. Beta-oksidasie
In die teenwoordigheid van lug en verskeie kofaktore en ensieme word vetsure na asetielkoënsiem-A omgeskakel. Die pad word beta-oksidasie genoem. Elke siklus van beta-oksidasie verkort die vetsuurketting met twee koolstofatome en produseer een ekwivalent elk van asetielkoënsiem-A, nikotinamiedadeniendinukleotied (NADH) en flavienadeniendinukleotied (FADH2). Die asetielkoënsiem-A word deur die sitroensuursiklus gemetaboliseer om ATP te genereer, terwyl die NADH en FADH2 deur oksidatiewe fosforilering gebruik word om ATP te genereer. Tientalle ATP-ekwivalente word gegenereer deur die beta-oksidasie van 'n enkele lang asielketting.[13]
Biochemiese funksies
Intrasellulêre seine
ATP is betrokke by seintransduksie deur as substraat vir kinase, ensieme wat fosfaatgroepe oordra, te dien.
DNS- en RNS-sintese
ATP is een van vier monomere wat nodig is in die sintese van RNS. Die proses word bevorder deur RNS-polimerase.[14] 'n Soortgelyke proses vind plaas in die vorming van DNS, behalwe dat ATP eers na die deoksiribonukleotied dATP omgeskakel word. Soos baie kondensasiereaksies in die natuur, verbruik DNS-replikasie en DNS-transkripsie ook ATP.
Aantekeninge
- 'n Hidrotroop is 'n verbinding wat hidrofobiese verbindings in waterige oplossings oplos op ander maniere as miselêre oplosbaarheid.
- 'n Nukleosiedtrifosfaat is 'n molekule wat 'n stikstofbasis bevat wat aan 'n 5-koolstof-suiker (óf ribose óf deoksiribose) gebind is, met drie fosfaatgroepe wat aan die suiker gebind is. Hulle dien as 'n bron van energie vir sellulêre reaksies.
- Die Krebs-siklus is 'n reeks chemiese reaksies om gestoorde energie vry te stel deur die oksidasie van asetielkoënsiem-A afkomstig van koolhidrate, vette en proteïene. Dit word ook die sitroensuursiklus genoem.
Verwysings
- O'Neil, Maryadele J. "Adenosine triphosphate". In Budavari, Susan (red.). The Merck index : an encyclopedia of chemicals, drugs, and biologicals (in Engels) (13de uitg.). Whitehouse Station, N.J. ISBN 0-911910-13-1. OCLC 52301949.
- Knowles, J.R. (1980). "Enzyme-catalyzed phosphoryl transfer reactions". Annu. Rev. Biochem. (in Engels). 49: 877–919. doi:10.1146/annurev.bi.49.070180.004305. PMID 6250450.
- Törnroth-Horsefield, S.; Neutze, R. (Desember 2008). "Opening and closing the metabolite gate". Proc. Natl. Acad. Sci. USA (in Engels). 105 (50): 19565–19566. Bibcode:2008PNAS..10519565T. doi:10.1073/pnas.0810654106. PMC 2604989. PMID 19073922.
- Storer, A.; Cornish-Bowden, A. (1976). "Concentration of MgATP2− and other ions in solution. Calculation of the true concentrations of species present in mixtures of associating ions". Biochem. J. (in Engels). 159 (1): 1–5. doi:10.1042/bj1590001. PMC 1164030. PMID 11772.
- Ferguson, S.J.; Nicholls, David; Ferguson, Stuart (2002). Bioenergetics 3 (in Engels) (3rd uitg.). San Diego, CA: Academic. ISBN 978-0-12-518121-1.
- Berg, J. M.; Tymoczko, J. L.; Stryer, L. (2003). Biochemistry (in Engels). New York, NY: W. H. Freeman. p. 376. ISBN 978-0-7167-4684-3.
- Chance, B.; Lees, H.; Postgate, J.G. (1972). "The Meaning of "Reversed Electron Flow" and "High Energy Electron" in Biochemistry". Nature (in Engels). 238 (5363): 330–331. Bibcode:1972Natur.238..330C. doi:10.1038/238330a0. PMID 4561837. S2CID 4298762.
- Beis, I.; Newsholme, E.A. (1 Oktober 1975). "The contents of adenine nucleotides, phosphagens and some glycolytic intermediates in resting muscles from vertebrates and invertebrates". Biochem. J. (in Engels). 152 (1): 23–32. doi:10.1042/bj1520023. PMC 1172435. PMID 1212224.
- Rich, P.R. (2003). "The molecular machinery of Keilin's respiratory chain". Biochem. Soc. Trans. (in Engels). 31 (6): 1095–1105. doi:10.1042/BST0311095. PMID 14641005.
- Lodish, H.; Berk, A.; Matsudaira, P.; Kaiser, C.A.; Krieger, M.; Scott, M.P.; Zipursky, S.L.; Darnell, J. (2004). Molecular Cell Biology (in Engels) (5th uitg.). New York, NY: W.H. Freeman. ISBN 978-0-7167-4366-8.
- Voet, D.; Voet, J.G. (2004). Biochemistry (in Engels). Vol. 1 (3de uitg.). Hoboken, NJ: Wiley. ISBN 978-0-471-19350-0.
- Schmidt-Rohr, K (2020). "Oxygen Is the High-Energy Molecule Powering Complex Multicellular Life: Fundamental Corrections to Traditional Bioenergetics". ACS Omega (in Engels). 5 (5): 2221–2233. doi:10.1021/acsomega.9b03352. PMC 7016920. PMID 32064383.
- Ronnett, G.; Kim, E.; Landree, L.; Tu, Y. (2005). "Fatty acid metabolism as a target for obesity treatment". Physiol. Behav. (in Engels). 85 (1): 25–35. doi:10.1016/j.physbeh.2005.04.014. PMID 15878185. S2CID 24865576.
- Joyce, C.M.; Steitz, T.A. (1995). "Polymerase structures and function: variations on a theme?". J. Bacteriol. (in Engels). 177 (22): 6321–6329. doi:10.1128/jb.177.22.6321-6329.1995. PMC 177480. PMID 7592405.